Sejtciklus: a G1-ből az S-fázisba való átmenet szabályozása. 21. §
A biológia érdekes és meglehetősen összetett témái közül érdemes kiemelni két sejtosztódási folyamatot a szervezetben - meiózis és mitózis. Elsőre úgy tűnhet, hogy ezek a folyamatok megegyeznek, hiszen mindkét esetben sejtosztódás történik, valójában azonban nagy különbség van köztük. Először is meg kell értened a mitózist. Mi ez a folyamat, mi a mitózis interfázisa és milyen szerepük van az emberi szervezetben? Erről ebben a cikkben részletesebben lesz szó.
Egy összetett biológiai folyamat, amelyet a sejtosztódás és a kromoszómák e sejtek közötti eloszlása kísér - mindez elmondható a mitózisról. Ennek köszönhetően a DNS-t tartalmazó kromoszómák egyenletesen oszlanak el a test leánysejtjei között.

A mitózis folyamatának 4 fő fázisa van. Mindegyik össze van kötve, mivel a fázisok zökkenőmentesen váltanak át egyikről a másikra. A természetben a mitózis elterjedtsége annak a ténynek köszönhető, hogy ez az, amely részt vesz az összes sejt osztódási folyamatában, beleértve az izmokat, az idegeket és így tovább.

Röviden az interfázisról
A mitózis állapotába kerülés előtt az osztódó sejt interfázisba kerül, azaz növekszik. Az interfázis időtartama normál üzemmódban a sejtaktivitás teljes idejének több mint 90%-át is elfoglalhatja.

Az interfázis 3 fő periódusra oszlik:
- fázis G1;
- S-fázis;
- fázis G2.
Mindegyik egy bizonyos sorrendben játszódik. Tekintsük ezeket a fázisokat külön-külön.

Interfázis – fő összetevők (képlet)
G1 fázis
Ezt az időszakot a sejt osztódásra való felkészítése jellemzi. A DNS-szintézis további fázisában növekszik a térfogata.
S-fázis
Ez az interfázis folyamat következő szakasza, amely során a szervezet sejtjei osztódnak. Általában a legtöbb sejt szintézise rövid idő alatt megy végbe. A sejtosztódás után a sejtek mérete nem nő, hanem az utolsó fázis kezdődik.

G2 fázis
Az interfázis utolsó szakasza, amelynek során a sejtek folytatják a fehérjék szintetizálását, miközben méretük nő. Ebben az időszakban még vannak sejtmagvak a sejtben. Ezenkívül az interfázis utolsó részében kromoszóma-duplikáció következik be, és a sejtmag felületét ebben az időben egy speciális héj borítja, amely védő funkcióval rendelkezik.
Egy megjegyzésben! A harmadik fázis végén mitózis lép fel. Több szakaszt is tartalmaz, amelyek után a sejtosztódás megtörténik (ezt a folyamatot az orvostudományban citokinézisnek nevezik).
A mitózis szakaszai
Amint azt korábban említettük, a mitózis 4 szakaszra oszlik, de néha több is lehet. Az alábbiakban a főbbeket.
Asztal. A mitózis főbb fázisainak ismertetése.
| Fázis neve, fotó | Leírás |
|---|---|
| A profázis során a kromoszómák spiralizálódnak, aminek következtében csavarodó alakot vesznek fel (tömörebb). A szervezet sejtjében minden szintetikus folyamat leáll, így a riboszómák már nem termelődnek. |
| Sok szakértő nem különbözteti meg a prometafázist a mitózis külön fázisaként. Gyakran az összes benne előforduló folyamatot profázisnak nevezik. Ebben az időszakban a citoplazma beburkolja a kromoszómákat, amelyek egy bizonyos pontig szabadon mozognak a sejtben. |
| A mitózis következő fázisa, amelyet a kondenzált kromoszómák egyenlítői síkon való eloszlása kísér. Ebben az időszakban a mikrotubulusok folyamatosan megújulnak. A metafázis során a kromoszómák úgy rendeződnek el, hogy a kinetochoreik eltérő irányúak, azaz ellentétes pólusok felé irányulnak. |
| A mitózis ezen fázisát az egyes kromoszómák kromatidjainak egymástól való elválasztása kíséri. A mikrotubulusok növekedése leáll, most kezdenek szétszedni. Az anafázis nem tart sokáig, de ezalatt az idő alatt a sejtek megközelítőleg egyenlő számban sikerül közelebb oszlani a különböző pólusokhoz. |
| Ez az utolsó szakasz, amely során a kromoszóma dekondenzációja megkezdődik. Az eukarióta sejtek befejezik osztódásukat, és az emberi kromoszómák mindegyike körül egy speciális héj képződik. Amikor a kontraktilis gyűrű összehúzódik, a citoplazma elválik (az orvostudományban ezt a folyamatot citotómiának nevezik). |





Fontos! A teljes mitózis folyamat időtartama általában nem haladja meg az 1,5-2 órát. Az időtartam a felosztandó cella típusától függően változhat. A folyamat időtartamát külső tényezők is befolyásolják, mint például a fényviszonyok, a hőmérséklet stb.

Milyen biológiai szerepe van a mitózisnak?
Most próbáljuk megérteni a mitózis jellemzőit és jelentőségét a biológiai ciklusban. Először is, biztosítja a szervezet számos létfontosságú folyamatát, beleértve az embrionális fejlődést is.

A mitózis a szervezet szöveteinek és belső szerveinek helyreállításáért is felelős különböző típusú károsodások után, ami regenerációt eredményez. A működés során a sejtek fokozatosan elpusztulnak, de a mitózis segítségével a szövetek szerkezeti integritása folyamatosan megmarad.

A mitózis bizonyos számú kromoszóma megőrzését biztosítja (az anyasejtben lévő kromoszómák számának felel meg).
Videó - A mitózis jellemzői és típusai
Ahhoz, hogy egy sejt teljesen osztódjon, meg kell nőnie a méretének és elegendő számú organellumnak kell lennie. És annak érdekében, hogy ne veszítse el az örökletes információkat, amikor kettéosztják, másolatot kell készítenie kromoszómáiról. És végül, annak érdekében, hogy az örökletes információkat szigorúan egyenlően ossza el két leánysejt között, a kromoszómákat a megfelelő sorrendbe kell rendeznie, mielőtt szétosztaná azokat a leánysejtek között. Mindezeket a fontos feladatokat a sejtciklus során hajtják végre.
A sejtciklus azért fontos, mert... a legfontosabbat mutatja be: a szaporodási, növekedési és differenciálódási képességet. Csere is előfordul, de ezt nem veszik figyelembe a sejtciklus tanulmányozása során.
A fogalom meghatározása
Sejtciklus - ez a sejt életének időszaka a születéstől a leánysejtek kialakulásáig.
Az állati sejtekben a sejtciklus, a két osztódás (mitózis) közötti időszak átlagosan 10-24 óráig tart.
A sejtciklus több periódusból (szinonimája: fázisokból) áll, amelyek természetesen helyettesítik egymást. A sejtciklus első fázisait (G 1, G 0, S és G 2) együttesen ún. interfázis , és az utolsó fázis neve .
Rizs. 1.Sejtciklus.
A sejtciklus periódusai (fázisai).
1. Az első növekedés időszaka G1 (az angol Growth - növekedésből) a ciklus 30-40%-a, a pihenőidő G. 0
Szinonimák: posztmitotikus (mitózis után következik be) periódus, preszintetikus (DNS szintézis előtt elhalad) időszak.
A sejtciklus a sejt mitózis útján történő születésével kezdődik. Az osztódás után a leánysejtek mérete csökken, és a normálisnál kevesebb organellumjuk van. Ezért egy „újszülött” kissejt a sejtciklus első periódusában (fázisában) (G 1) növekszik és megnövekszik, és kialakítja a hiányzó organellumokat is. Mindehhez szükséges a fehérjék aktív szintézise. Ennek eredményeként a sejt teljes értékűvé válik, mondhatni „felnőttté”.
Hogyan végződik a G1 növekedési periódus általában egy sejt esetében?
- A sejt belépése a folyamatba. A differenciálódásnak köszönhetően a sejt különleges tulajdonságokat szerez az egész szerv és szervezet számára szükséges funkciók ellátásához. A differenciálódást a sejt megfelelő molekuláris receptoraira ható szabályozó anyagok (hormonok) váltják ki. A differenciálódást befejező sejt kiesik az osztódási ciklusból, és benne van pihenőidő G 0 . Aktiváló anyagokkal (mitogénekkel) való érintkezés szükséges ahhoz, hogy dedifferenciálódjon és visszatérjen a sejtciklusba.
- A sejt halála (halála).
- Belépés a sejtciklus következő időszakába - szintetikus.
2. Az S szintetikus periódus (az angol Synthesis - szintézis szóból) a ciklus 30-50%-át teszi ki
A szintézis fogalma e korszak nevében arra utal DNS-szintézis (replikáció) , és nem más szintézis folyamatokhoz. Miután az első növekedési perióduson áthaladva elért egy bizonyos méretet, a sejt belép a szintetikus periódusba, vagy az S fázisba, amelyben a DNS-szintézis megtörténik. A DNS replikációja miatt a sejt megkétszerezi genetikai anyagát (kromoszómák), mert Mindegyik kromoszóma pontos másolata keletkezik a sejtmagban. Minden kromoszóma megkettőződik, és a teljes kromoszómakészlet kettős lesz, ill diploid . Ennek eredményeként a sejt készen áll arra, hogy az örökítőanyagot egyenlő arányban megosszák két leánysejt között anélkül, hogy egyetlen gént is elveszítenének.
3. A második növekedés időszaka G 2 (az angol Growth - növekedésből) a ciklus 10-20%-a
Szinonimák: premitotikus (a mitózis előtt elmúlik) időszak, posztszintetikus (a szintetikus után következik be) időszak.
A G2 periódus előkészíti a következő sejtosztódást. A G2 növekedés második periódusában a sejt a mitózishoz szükséges fehérjéket, különösen az orsóhoz szükséges tubulint termel; energiatartalékokat hoz létre ATP formájában; ellenőrzi, hogy a DNS-replikáció befejeződött-e, és felkészül az osztódásra.
4. Az M mitózisos osztódás időszaka (az angol Mitosis - mitosis szóból) a ciklus 5-10%-a
Az osztódás után a sejt új G1 fázisba lép, és a sejtciklus véget ér.


Sejtciklus szabályozás
Molekuláris szinten a ciklus egyik fázisából a másikba való átmenetet két fehérje szabályozza - ciklinÉs ciklin-függő kináz(CDK).
A sejtciklus szabályozására a szabályozó fehérjék reverzibilis foszforilációjának/defoszforilációjának folyamatát alkalmazzák, azaz. foszfátok hozzáadása, majd elimináció. A sejt mitózisba való belépését (azaz a G2 fázisból az M fázisba való átmenetét) szabályozó kulcsanyag egy specifikus szerin/treonin protein kináz, ami az úgynevezett érési tényező- FS, vagy MPF, az angol érést elősegítő faktorból. Aktív formájában ez a fehérje enzim katalizálja a mitózisban részt vevő számos fehérje foszforilációját. Ilyen például a H1 hiszton, amely a kromatin része, a lamin (a sejtmagmembránban található citoszkeletális komponens), a transzkripciós faktorok, a mitotikus orsófehérjék, valamint számos enzim. Ezeknek a fehérjéknek az MPF érési faktor általi foszforilációja aktiválja őket, és beindítja a mitózis folyamatát. A mitózis befejezése után a PS szabályozó alegység, ciklin, ubiquitinnel van megjelölve, és lebomlik (proteolízis). Most rajtad a sor protein foszfatáz, amelyek defoszforilálják a mitózisban részt vevő fehérjéket, ezáltal inaktív állapotba kerülnek. Ennek eredményeként a sejt visszatér az interfázis állapotba.
A PS (MPF) egy heterodimer enzim, amely tartalmaz egy szabályozó alegységet, nevezetesen a ciklint, és egy katalitikus alegységet, nevezetesen a ciklinfüggő kináz CDK-t, más néven p34cdc2; 34 kDa. Ennek az enzimnek az aktív formája csak a CZK + ciklin dimer. Ezenkívül a CZK aktivitást magának az enzimnek a reverzibilis foszforilációja szabályozza. A ciklinek azért kapták ezt a nevet, mert koncentrációjuk ciklikusan változik a sejtciklus periódusainak megfelelően, különösen a sejtosztódás kezdete előtt csökken.
Számos különböző ciklin és ciklinfüggő kináz található a gerinces sejtekben. Két enzimalegység különböző kombinációi szabályozzák a mitózis beindulását, a transzkripciós folyamat kezdetét a G1 fázisban, a kritikus pont átmenetét a transzkripció befejezése után, a DNS replikációs folyamatának kezdetét az interfázis S periódusában (átmenet kezdete). ) és a sejtciklus egyéb kulcsátmenetei (az ábrán nem látható).
A béka petesejtekben a mitózisba való belépést (G2/M átmenet) a ciklinkoncentráció változásai szabályozzák. A ciklin az interfázisban folyamatosan szintetizálódik, amíg az M fázisban el nem éri a maximális koncentrációt, amikor is beindul a PS által katalizált fehérjefoszforiláció teljes kaszkádja. A mitózis végére a ciklint a PS által is aktivált proteinázok gyorsan elpusztítják. Más sejtrendszerekben a PS-aktivitást magának az enzimnek a foszforilációjának különböző mértéke szabályozza.
Sejtciklus
A sejtciklus mitózisból (M fázis) és interfázisból áll. Az interfázisban a G 1, S és G 2 fázisokat egymás után megkülönböztetjük.
A SEJTCIKLUS SZAKASZAI
Interfázis
G 1 követi a mitózis telofázisát. Ebben a fázisban a sejt RNS-t és fehérjéket szintetizál. A fázis időtartama több órától több napig tart.
G 2 A sejtek kiléphetnek a ciklusból és fázisban vannak G 0 . fázisban G 0 a sejtek elkezdenek differenciálódni.
S. Az S-fázisban a fehérjeszintézis folytatódik a sejtben, DNS-replikáció következik be, és a centriolok szétválnak. A legtöbb sejtben az S fázis 8-12 óráig tart.
G 2 . A G 2 fázisban az RNS és a fehérje szintézise folytatódik (például a tubulin szintézise a mitotikus orsó mikrotubulusaihoz). A leány centriolák elérik a definitív organellumok méretét. Ez a fázis 2-4 óráig tart.
MITÓZIS
A mitózis során a mag (kariokinézis) és a citoplazma (citokinézis) osztódik. A mitózis fázisai: profázis, prometafázis, metafázis, anafázis, telofázis.
Prophase. Mindegyik kromoszóma két testvérkromatidból áll, amelyeket egy centromer köt össze, a mag eltűnik. A centriolesok szervezik a mitotikus orsót. A mitotikus központ része egy centriolpár, amelyből a mikrotubulusok sugárirányban nyúlnak ki. Először a mitotikus centrumok a nukleáris membrán közelében helyezkednek el, majd szétválnak, és egy bipoláris mitotikus orsó képződik. Ez a folyamat pólusmikrotubulusokat foglal magában, amelyek megnyúlásuk során kölcsönhatásba lépnek egymással.
Centriole része a centroszómának (a centroszóma két centriolt és egy pericentriolmátrixot tartalmaz), és 15 nm átmérőjű és 500 nm hosszú henger alakú; a hengerfal 9 mikrotubulus hármasból áll. A centroszómában a centriolák egymásra merőlegesen helyezkednek el. A sejtciklus S fázisában a centriolok megkettőződnek. A mitózisban az eredeti és egy újonnan képződött centriolák párjai a sejtpólusokhoz térnek el, és részt vesznek a mitotikus orsó kialakításában.
Prometafázis. A nukleáris burok apró darabokra bomlik. A centromerek régiójában kinetokorok jelennek meg, amelyek a kinetochore mikrotubulusok szerveződésének központjaként működnek. A kromoszómák mozgásának oka a kinetokorok mindkét irányban történő távozása az egyes kromoszómákból, valamint a mitotikus orsó pólusmikrotubulusaival való kölcsönhatásuk.
Metafázis. A kromoszómák az orsó egyenlítői régiójában helyezkednek el. Metafázis lemez képződik, amelyben minden kromoszómát egy pár kinetochore és a kapcsolódó kinetochore mikrotubulusok tartanak, amelyek a mitotikus orsó ellentétes pólusaira irányulnak.
Anafázis– a leánykromoszómák divergenciája a mitotikus orsó pólusaihoz 1 µm/perc sebességgel.
Telofázis. A kromatidák megközelítik a pólusokat, a kinetochore mikrotubulusok eltűnnek, a pólusok pedig tovább nyúlnak. Kialakul a magburok, és megjelenik a mag.
Citokinézis– a citoplazma szétválása két különálló részre. A folyamat késői anafázisban vagy telofázisban kezdődik. A plazmalemma visszahúzódik a két leánymag közé az orsó hossztengelyére merőleges síkban. A hasítási barázda mélyül, és a leánysejtek között híd marad - egy maradék test. Ennek a szerkezetnek a további megsemmisítése a leánysejtek teljes elválasztásához vezet.
A sejtosztódás szabályozói
A sejtproliferációt, amely mitózison keresztül megy végbe, számos molekuláris jel szigorúan szabályozza. Ezeknek a többszörös sejtciklus-szabályozóknak az összehangolt tevékenysége biztosítja a sejtek átmenetét a sejtciklus egyik fázisáról a másikra, valamint az egyes fázisok eseményeinek pontos végrehajtását. A proliferatívan kontrollálatlan sejtek megjelenésének fő oka a sejtciklus-szabályozók szerkezetét kódoló gének mutációi. A sejtciklus és a mitózis szabályozói intracellulárisra és intercellulárisra oszthatók. Az intracelluláris molekuláris szignálok számosak, ezek közül mindenekelőtt maguk a sejtciklus-szabályozók (ciklinek, ciklin-dependens protein kinázok, ezek aktivátorai és inhibitorai) és a tumorszuppresszorok említendők.
MEIOSIS
A meiózis során haploid ivarsejtek képződnek.
Első meiotikus osztódás
A meiózis első felosztása (I. profázis, I. metafázis, I. anafázis és I. telofázis) a redukció.
Prophaseén egymás után több szakaszon megy keresztül (leptoten, zigotén, pachitén, diplotén, diakinézis).
Leptoten – A kromatin kondenzálódik, minden kromoszóma két kromatidából áll, amelyeket centromer köt össze.
Zigotén– a homológ páros kromoszómák közelebb kerülnek egymáshoz és fizikai érintkezésbe kerülnek ( szinapszis) szinaptonemális komplex formájában, amely biztosítja a kromoszómák konjugációját. Ebben a szakaszban két szomszédos kromoszómapár alkot egy bivalenst.
Pachytena– a kromoszómák megvastagodnak a spiralizáció miatt. A konjugált kromoszómák külön szakaszai metszik egymást, és chiasmát alkotnak. Itt történik átkelés- metszetcsere az apai és anyai homológ kromoszómák között.
Diplotena– a konjugált kromoszómák szétválása minden párban a synaptonemalis komplex longitudinális hasadása következtében. A kromoszómák a komplex teljes hosszában hasadnak, a chiasmata kivételével. A bivalensben 4 kromatid jól megkülönböztethető. Az ilyen bivalenst tetradnak nevezik. A kromatidákban letekeredő helyek jelennek meg, ahol az RNS szintetizálódik.
Diakinézis. Folytatódnak a kromoszóma rövidülésének és a kromoszómapárok szétválásának folyamatai. A chiasma a kromoszómák végére költözik (terminalizáció). A nukleáris membrán megsemmisül, és a sejtmag eltűnik. Megjelenik a mitotikus orsó.
Metafázisén. Az I. metafázisban a tetradák alkotják a metafázis lemezt. Általában az apai és anyai kromoszómák véletlenszerűen oszlanak el a mitotikus orsó egyenlítőjének egyik vagy másik oldalán. Ez a kromoszómaeloszlási mintázat alapozza meg Mendel második törvényét, amely (a keresztezéssel együtt) biztosítja az egyedek közötti genetikai különbségeket.
Anafázisén abban különbözik a mitózis anafázisától, hogy a mitózis során a testvérkromatidák a pólusok felé mozognak. A meiózis ezen fázisában az ép kromoszómák a pólusokra költöznek.
Telofázisén nem különbözik a mitózis telofázisától. 23 konjugált (kettős) kromoszómát tartalmazó magok képződnek, citokinézis lép fel, és leánysejtek képződnek.
A meiózis második osztódása.
A meiózis második felosztása - egyenletek - ugyanúgy megy végbe, mint a mitózis (profázis II, metafázis II, anafázis II és telofázis), de sokkal gyorsabban. A leánysejtek egy haploid kromoszómakészletet kapnak (22 autoszóma és egy nemi kromoszóma).
GOUVPO
"VORONEZI ÁLLAMI MŰSZAKI EGYETEM"
ORVOSI RENDSZEREK RENDSZERELEMZÉSI ÉS -MENEDZSMENTI OSZTÁLYA
ABSZTRAKT
Fegyelem: „Ember- és állatbiológia”
A TÉMÁRÓL: „Mitotikus ciklus. Sejtciklus, M, G1, S, G2 fázisok, auto- és heteroszintetikus sejtfunkciók"
Elkészítette: a BM-101 csoport 1. éves hallgatója Tonkikh M.A.
Ellenőrizte: professzor, orvosdoktor. Tudomány L. B. Dmitrenko
VORONEZH 2010
Sejtciklus: áttekintés
Az eukarióta sejtek osztódását biztosító ismétlődő események sorozatát sejtciklusnak nevezzük. A sejtciklus hossza az osztódó sejtek típusától függ. Egyes sejtek, például az emberi neuronok, teljesen leállnak az osztódással, miután elérik a terminális differenciálódás szakaszát. A tüdő, a vese vagy a máj sejtjei egy felnőtt szervezetben csak a megfelelő szervek károsodására reagálva kezdenek osztódni. A bélhámsejtek az ember élete során osztódnak. A gyorsan szaporodó sejtekben is körülbelül 24 órát vesz igénybe az osztódásra való felkészülés. A sejtciklus szakaszokra oszlik: Mitózis - M-fázis, a sejtmag osztódása. A G1 fázis a DNS szintézis előtti időszak. Az S-fázis a szintézis (DNS-replikáció) periódusa. A G2 fázis a DNS-szintézis és a mitózis közötti időszak. Az interfázis egy olyan időszak, amely magában foglalja a G1, S és G2 fázisokat. A citokinézis a citoplazma osztódása. Korlátozási pont, R-pont – az az idő a sejtciklusban, amikor a sejt osztódása felé visszafordíthatatlanná válik. A G0-fázis azon sejtek állapota, amelyek a korai G1-fázisban elértek egyrétegűt vagy megfosztják a növekedési faktoroktól.
mitozumeiosis) kromoszóma megkettőződése előzi meg, amely a sejtciklus S periódusában következik be. Az időszakot a szintézis szó első betűje jelöli - DNS-szintézis. Az S periódus végétől a metafázis végéig a sejtmag négyszer több DNS-t tartalmaz, mint a spermium vagy a tojás magja, és minden kromoszóma két azonos testvérkromatidából áll.
A mitózis során a kromoszómák kondenzálódnak és a profázis végén vagy a metafázis elején optikai mikroszkóppal láthatóvá válnak. A citogenetikai elemzéshez általában metafázisú kromoszómák preparátumait használják.
Először homológ kromoszómák anafázis centroméra le vannak kapcsolva, és kromatidák eltérnek a mitotikus orsó ellentétes pólusaihoz. Miután a kromatidák teljes készletei a pólusokra költöznek (a továbbiakban kromoszómáknak hívják őket), mindegyik körül egy magmembrán képződik, amely két leánysejt magját képezi (az anyasejt magmembránjának pusztulása vége próféta). Leánysejtek lépnek be időszak G1, és csak a következő osztódásra készülve lépnek be az S periódusba és DNS-replikáció megy végbe bennük. 
Azok a speciális funkciójú sejtek, amelyek hosszú ideig nem lépnek be mitózisba, vagy általában elvesztették osztódási képességüket, ún. időszak G0 .
A legtöbb sejt a testben diploid – vagyis kettő van haploid kromoszómakészlet(a haploid halmaz az ivarsejtekben lévő kromoszómák száma; emberben 23 kromoszóma, ill. diploid kromoszómakészlet - 46).
Az ivarmirigyekben a csírasejtek prekurzorai először egy sor mitotikus osztódáson mennek keresztül, majd belépnek a meiózisba, az ivarsejtek képződésének folyamatába, amely két egymást követő osztódásból áll. A meiózisban homológ kromoszómák párosulnak (apai 1. kromoszóma az anyai 1. kromoszómával stb.), ami után az ún. átkelés rekombináció történik, vagyis szakaszok cseréje az apai és az anyai kromoszómák között. Ennek eredményeként az egyes kromoszómák genetikai összetétele minőségileg megváltozik.
Az első osztályban meiózis Homológ kromoszómák (és nem testvérkromatidák, mint pl mitózis), amelynek eredményeként haploid kromoszómakészlettel rendelkező sejtek képződnek, amelyek mindegyike 22 megkétszerezett kromoszómát tartalmaz. autoszómákés egy megkettőzött nemi kromoszóma.
A meiózis első és második osztódása között nincs S időszak ( rizs. 66.2, jobbra), és a testvérkromatidák leánysejtekké válnak szét a második osztódásban. Ennek eredményeként haploid kromoszómakészlettel rendelkező sejtek képződnek, amelyekben a G1 periódusban feleannyi DNS van, mint a diploid szomatikus sejtekben, és 4-szer kevesebb, mint az S periódus végén.
A megtermékenyítés során a zigótában a kromoszómák száma és a DNS-tartalom megegyezik a G1 periódusban lévő szomatikus sejttel.
Az S periódus a zigótában megnyitja az utat a szomatikus sejtekre jellemző szabályos osztódás felé.
Sejtciklus: fázisok
Az eukarióta sejtciklus négy fázisra oszlik. A közvetlen sejtosztódás (mitózis) szakaszában a kondenzált metafázisú kromoszómák egyenlően oszlanak el a leánysejtek között. A sejtciklus M fázisa - mitózis). A mitózis volt az azonosított sejtciklus első fázisa, és a sejtben két mitózis között előforduló összes többi eseményt ún. interfázis. A molekuláris szintű kutatás fejlődése lehetővé tette a DNS-szintézis interfázisban lévő szakaszának, ún. S-fázis (szintézis). A sejtciklus e két kulcsfontosságú szakasza nem megy át közvetlenül egymásba. A mitózis vége után, mielőtt a DNS-szintézis megkezdődik, A sejtciklus G1 fázisa (rés), a sejtaktivitás látszólagos szünete, amely során az intracelluláris szintetikus folyamatok előkészítik a genetikai anyag replikációját.
Második szünet a látható tevékenységben ( fázis G2) a DNS-szintézis befejezése után, a mitózis kezdete előtt figyelhető meg. A G2 fázisban a sejt figyeli a megtörtént DNS-replikáció pontosságát, és kijavítja az észlelt hibákat. Egyes esetekben a sejtciklus ötödik fázisa megkülönböztethető ( G0), amikor az osztódás befejezése után a sejt nem lép be a következő sejtciklusba, és hosszú ideig alvó állapotban marad. Ebből az állapotból külső stimuláló (mitogén) hatások hatására eltávolítható.
A sejtciklus fázisai nem rendelkeznek egyértelmű időbeli és funkcionális határokkal, azonban az egyik fázisból a másikba való átmenet során a szintetikus folyamatok rendezett átváltása történik, ami lehetővé teszi ezen intracelluláris események molekuláris szintű differenciálódását.
Ciklinek és ciklin-dependens kinázok
A sejtek belépnek a sejtciklusba, és külső mitogén ingerekre válaszul DNS-t szintetizálnak. Limfokinok(Például, interleukinok), citokinek(különösen interferonok) és a polipeptid növekedési faktorok a sejtfelszínen lévő receptoraikkal kölcsönhatásba lépve intracelluláris fehérjék foszforilációs reakcióinak sorozatát indukálják, amelyet jelátvitel kísér a sejtfelszínről a sejtmagba és a megfelelő gének transzkripciójának indukciója. Az elsők között aktiválódnak a kódoló gének ciklin fehérjék, amelyek nevüket onnan kapták, hogy intracelluláris koncentrációjuk időszakosan változik, ahogy a sejtek áthaladnak a sejtcikluson, és bizonyos szakaszokban elérik a maximumot. A ciklinek a család sajátos aktivátorai ciklinfüggő protein kinázok (CDK) (CDK - ciklin-dependens kinázok) kulcsfontosságú résztvevői a sejtciklust szabályozó gének transzkripciójának indukciójában. Egy egyedi CDK aktiválódása egy adott ciklinnal való kölcsönhatás után következik be, és ennek a komplexnek a kialakulása akkor válik lehetővé, amikor a ciklin eléri a kritikus koncentrációt. Egy adott ciklin intracelluláris koncentrációjának csökkenésére adott válaszként a megfelelő CDK reverzibilisen inaktiválódik. Néhány CDK-t egynél több ciklin aktivál. Ebben az esetben a ciklinek egy csoportja, mintha proteinkinázokat adna át egymásnak, hosszú ideig aktivált állapotban tartja őket. A CDK aktiválódásának ilyen hullámai a sejtciklus G1 és S fázisában fordulnak elő.
Ciklinek: általános információk
Minden A-tól H-ig jelölő ciklin típusnak van egy homológ régiója (150 aminosav, az úgynevezett " ciklin doboz". Ez a régió felelős a kötődésért CDK. A ciklin családban 14 fehérje ismert (ciklin A - ciklin J). A család egyes tagjai alcsaládokat alkotnak. Például a D-típusú ciklin alcsalád három tagból áll: a D1, D2 és D3 ciklineket két alcsaládra osztják: G1-ciklinek (C , DÉs E) És mitotikus ciklinek (AÉs B).
A ciklinek gyorsan cserélik a fehérjéket, rövid felezési idővel, ami a D-típusú ciklinek esetében 15-20 perc. Ez biztosítja a komplexek dinamizmusát ciklinfüggő kinázok. Az aminosavak N-terminális szekvenciáját ún megsemmisítő doboz. Ahogy a sejtek áthaladnak a sejtcikluson, az egyed aktiválódását követően CDK szükség szerint inaktiválva vannak. Ez utóbbi esetben a CDK-val komplexben lévő ciklin proteolitikus lebomlása megy végbe, ami egy destrukciós dobozzal kezdődik.
A ciklinek maguk nem tudják teljesen aktiválni a megfelelő CDK-kat. Az aktiválási folyamat befejezéséhez ezeknek a protein-kinázoknak a polipeptidláncában bizonyos aminosavak specifikus foszforilációjának és defoszforilációjának meg kell történnie. A legtöbb ilyen reakciót végrehajtják CDK-aktiváló kináz (CAK), ami egy komplex CDK7 Val vel ciklin H. Így a CDK-k csak a megfelelő ciklinekkel való kölcsönhatás és a CAK és más hasonló sejtciklust szabályozó fehérjék hatására bekövetkező poszttranszlációs módosulások után válnak alkalmassá a sejtciklusban betöltött funkcióik ellátására.
Eukarióta sejtosztódás: kezdet
Mitogén ingerre adott válaszként egy sejt be fázis G0 vagy korán G1, megkezdi áthaladását a sejtcikluson. Az expresszió indukciójának eredményeként ciklin D génekÉs E, amelyeket általában csoportosítanak ciklinek G1, növekszik az intracelluláris koncentrációjuk. Ciklinek D1 , D2És D3 komplexet alkotnak a kinázokkal CDK4És CDK6. A ciklin D1-től eltérően az utóbbi két ciklin is kombinálódik CDK2. A három ciklin funkcionális különbségei nem ismertek, de a rendelkezésre álló adatok azt mutatják, hogy a G1 fázis fejlődésének különböző szakaszaiban érik el a kritikus koncentrációt. Ezek a különbségek a proliferáló sejtek típusára jellemzőek.
A CDK2/4/6 aktiválása foszforilációhoz vezet mókus RB(termék retinoblasztóma gén pRb) és a kapcsolódó fehérjék 107. oÉs 130. o. A G1 fázis elején pRb fehérje gyengén foszforilált, ami lehetővé teszi, hogy komplexben legyen vele transzkripciós faktor E2F, amely kulcsszerepet játszik a DNS szintézis indukálásában, és gátolja annak aktivitását. A pRb teljesen foszforilált formája E2F-et szabadít fel a komplexből, ami a DNS-replikációt szabályozó gének transzkripciós aktiválásához vezet.
A D-ciklinek koncentrációja a sejtciklus G1 fázisában növekszik, és közvetlenül a sejtciklus kezdete előtt éri el maximális értékét. S-fázis, utána csökkenni kezd. Azonban ebben az időben a pRb még nem foszforilálódott teljesen, és az E2F faktor inaktív állapotban marad a komplexben. A pRb foszforilációja a CDK2 aktiválásával fejeződik be ciklin E. Ez utóbbi intracelluláris koncentrációja a sejtciklus G1 fázisból S fázisba való átmenetének pillanatában válik maximálissá. Így a ciklin E-CDK2 komplex átveszi a ciklin D komplexeket CDK4-gyel és CDK6-tal, és befejezi a pRb foszforilációját, amelyet az E2F aktív transzkripciós faktor felszabadulása kísér. Ennek hatására megindul a DNS-szintézis, vagyis a sejt belép a sejtciklus S-fázisába.
A sejtciklus S fázisa: DNS szintézis
Időszak interfázis amikor a sejtmag DNS-replikációja megtörténik, "S-fázisnak" nevezték.
A sejtosztódást (mitózist vagy meiózist) kromoszóma-duplikáció előzi meg, ami a sejtciklus S periódusában ( rizs. 66.2). Az időszakot a szintézis szó első betűje jelöli - DNS-szintézis.
Miután a sejt az S fázisba lép, gyors lebomlás következik be ciklin Eés aktiválás CDK2 ciklin A. A Cyclin E szintetizálása a végén kezdődik fázis G1és a CDK2-vel való kölcsönhatása szükséges feltétele annak, hogy a sejt S-fázisba lépjen, és folytassa a DNS-szintézist. Ez a komplex aktiválja a DNS-szintézist a fehérjék foszforilációjával a replikációs origónál. Jel az S-fázis végére és a cella átmenetére fázis G2 egy másik kináz aktiválása ciklin A által CDK1 a CDK2 aktiválásának egyidejű leállításával. Késés a DNS-szintézis vége és a kezdete között mitózis(G2 fázis) a sejt által a megtörtént kromoszóma replikáció teljességének és pontosságának szabályozására szolgál. Az eseménysorozat ebben az időszakban nem ismert pontosan.
Amikor stimulálják növekedési tényezők-ben található emlőssejtek proliferatív nyugalmi állapot , ciklinek D-típusúak korábban jelennek meg, mint a ciklin E. mRNS és fehérje ciklin D1 először 6-8 óra elteltével jelennek meg, ezt követően a D1 szint a sejtciklus végéig emelkedett marad ( Matsushime H. et al., 1991 ; Megnyerte a K.A. et al., 1992).
Amikor a növekedési faktorokat eltávolítjuk a tápközegből, a D-típusú ciklinek szintje gyorsan csökken, mivel a D-ciklinek és RNS-ük instabil.
Ciklin D1 társult, összekapcsolt, társított valamivel CDK4 közvetlenül a DNS-szintézis megkezdése előtt. A komplex szintje a korai S fázisban tetőzik, mielőtt csökken a késői S és G2 fázis (Matsushime H. et al., 1992).
Látszólag ciklinek D2És D3 a G1 periódusban valamivel később hat, mint a D1 ciklin.
A D-típusú ciklinek túlzott expressziója (a normálhoz képest ötszöröse) a növekedési faktorok iránti sejtek iránti igény csökkenésével és a G1 fázis lerövidülésével a sejtméret csökkenéséhez vezet. Ciklin E szükséges a sejtek bejutásához S-fázis. Elsősorban azzal társul CDK2, bár komplexet képezhet vele CDK1 .
A ciklin E mRNS és fehérje szintje, valamint a ciklin E-CDK2 komplex aktivitása az átmenet során tetőzik G1-Sés élesen csökken, ahogy a sejtek előrehaladnak a középső és késői S fázison.
Ha ciklin E elleni antitesteket mikroinjektálnak emlőssejtekbe, a DNS szintézis elnyomódik.
Ha a ciklin E túlzottan expresszálódik, a sejtek gyorsabban haladnak át a G1 fázison, és belépnek az S fázisba, és az ilyen sejtek kevesebb növekedési faktort igényelnek.
Mitózis: beindítás
A sejtosztódás (mitózis) megindulásának jele innen származik MPF faktor (M fázist elősegítő faktor), stimulálja a sejtciklus M fázisát. Az MPF egy kináz komplex CDK1 aktiválásával ciklinek A vagy B. Úgy tűnik, a CDK1-ciklin A komplex fontosabb szerepet játszik az S fázis befejezésében és a sejt osztódásra való felkészítésében, míg a CDK1-ciklin B komplex elsősorban a szekvencia szabályozását gyakorolja.
Ciklinek B1És B2 nagyon alacsony koncentrációban van jelen fázis G1. Koncentrációjuk a vége felé növekedni kezd S-és az egész G2 fázisok, a mitózis során érik el maximumát, ami pótlásukhoz vezet ciklin A kombinálva valamivel CDK1. Ez azonban nem elegendő a protein-kináz teljes aktiválásához. A CDK1 funkcionális kompetenciája egy sor foszforiláció és defoszforiláció után érhető el specifikus aminosavaknál. Az ilyen szabályozásra azért van szükség, hogy megakadályozzuk a sejtek mitózisba jutását, amíg a DNS-szintézis be nem fejeződik.
A sejtosztódás csak azután kezdődik, hogy a CDK1, amely a ciklin B-vel komplexben van, a Thr-14 és Tyr-16 aminosavaknál foszforilálódik protein kináz WEE1, valamint a Thr-161 maradéknál protein kináz CAK majd a Thr-14 és Tyr-15 maradékoknál defoszforilálják foszfatáz CDC25. Ily módon aktiválva a CDK1 foszforilálja a sejtmagban lévő strukturális fehérjéket, beleértve nukleolin , nukleáris laminákÉs vimentin. Ezt követően a sejtmag elkezd áthaladni a mitózis citológiailag egyértelműen megkülönböztethető szakaszain.
A mitózis első szakasza az próféta- után kezdődik CDK1 teljesen foszforilálódik, ezt követi metafázis , anafázisÉs telofázis sejtosztódással végződve citokinézis. Ezeknek a folyamatoknak a következménye a replikált kromoszómák, nukleáris és citoplazmatikus fehérjék, valamint egyéb nagy és kis molekulatömegű vegyületek megfelelő eloszlása a leánysejtekbe. A citokinézis befejeződése után pusztulás következik be ciklin B, amit a CDK1 inaktiválódása kísér, ami a sejt bejutásához vezet fázis G1 vagy G0 sejtciklus.
A sejtciklus G0 fázisa
Egyes sejttípusok a differenciálódás bizonyos szakaszaiban leállíthatják az osztódást, teljes mértékben fenntartva életképességüket. A sejteknek ezt az állapotát G0 fázisnak nevezik. Azok a sejtek, amelyek elérték a terminális differenciálódás állapotát, már nem tudnak kilépni ebből a fázisból. Ugyanakkor a rendkívül alacsony osztódási képességű sejtek, például a hepatociták, a máj egy részének eltávolítása után újra beléphetnek a sejtciklusba.
A sejtek nyugalmi állapotba való átmenete a rendkívül specifikus működésének köszönhetően válik lehetővé sejtciklus-gátlók. Ezeknek a fehérjéknek a részvételével a sejtek leállíthatják a proliferációt kedvezőtlen környezeti feltételek mellett, amikor a DNS károsodik, vagy a replikációjában durva hibák lépnek fel. Az ilyen szüneteket a sejtek a bekövetkezett károsodások helyreállítására használják.
Bizonyos külső körülmények között a sejtciklus megszakadhat korlátozási pontok. Ezeken a pontokon a sejtek elköteleződnek az S-fázisba és/vagy a mitózisba való belépésre.
Gerinces sejtek szabványos táptalajban, amely mentes szérum, A legtöbb esetben ne lépjen be az S-fázisba, bár a táptalaj minden szükséges tápanyagot tartalmaz.
A zárt egyrétegű réteg elérésekor a sejtek képesek érintkező fékezés, jelenlétében is kilép a sejtciklusból vérszérum. Azokat a sejteket, amelyek határozatlan időre elhagyták a mitotikus ciklust, fenntartva az életképességet és a proliferációs potenciált, nyugalmi sejteknek nevezzük. Ezt a proliferatív nyugalmi állapotba vagy a G0 fázisba való átmenetnek nevezik.
A 90-es években Folytatódtak a viták, hogy a proliferatív nyugalmi állapot a G1-től alapvetően eltérő fázisként definiálható-e. Nyilvánvalóan ez a helyzet.
A proliferatív nyugalomban lévő sejtek magjaiban, valamint azokban a sejtekben, amelyek a G1 fázis, rendszerint kétségtelenül sok DNS-t tartalmaz. Ebben a két állapotban azonban jelentős különbségek vannak a sejtek között. Ismeretes, hogy az osztódó sejtekben a G1 fázis időtartama lényegesen rövidebb, mint a G0-S átmenet ideje. A nyugvó és proliferáló sejtek fúziójával és az mRNS mikroinjekciójával kapcsolatos számos tanulmány kimutatta, hogy a G0 fázisban lévő sejtek tartalmaznak proliferáció gátlók, megakadályozva az S-fázisba való belépést.
Ezek a tények arra utalnak, hogy a cellának egy speciális programot kell végrehajtania a G0-ból való kilépéshez. Azt is meg kell jegyezni, hogy nyugalmi sejtekben nem expresszálódnak. CDK2És CDK4, és ciklinek D- És E-típusok. Szintézisüket csak növekedési faktorok indukálják ( Lodish H. et al., 1995). BAN BEN folyamatosan ciklikus sejtek a D- és E-ciklinek szintje a teljes ciklus alatt magas marad, és a G1 periódus időtartama csökken a prereplikációs periódushoz képest.
Így a G0 fázis sejtjeiben nincs olyan fehérje, amely lehetővé tenné a restrikciós pontokon való átjutást és az S fázisba való bejutást. A nyugvó sejtek S-fázisba való átmenetéhez növekedési tényezők ezeknek a fehérjéknek a szintézisét kell indukálnia bennük.
Sejtciklus: inhibitorok
A sejtciklusnak két fő szakasza van (átmeneti pontok, ellenőrzési pontok R - korlátozási pontok), amelyeken megvalósíthatók negatív szabályozási hatások, megakadályozza a sejtek mozgását a sejtcikluson. Ezen szakaszok egyike a sejt DNS-szintézisbe való átmenetét, a másik pedig a mitózis kezdetét szabályozza. A sejtciklusnak vannak más szabályozott szakaszai is.
A sejtek átmenetét a sejtciklus egyik fázisából a másikba az aktiválás szintjén szabályozzák CDK az övék ciklinek val vel ciklin-dependens kinázok inhibitorai, CKI. Szükség esetén ezek az inhibitorok aktiválhatók, és blokkolják a CDK-k kölcsönhatását ciklineikkel, és ezáltal magát a sejtciklust. A külső vagy belső körülmények megváltozása után a sejt tovább szaporodhat, vagy beléphet az útba apoptózis .
Két CKI-csoport létezik: p21 család fehérjéiÉs INK4 (CDK4 inhibitor), amelyek családon belüli tagjai hasonló szerkezeti tulajdonságokkal rendelkeznek. A p21 inhibitorcsalád három fehérjét tartalmaz: p21 , 27. oÉs 57. o. Mivel ezeket a fehérjéket több csoport egymástól függetlenül írta le, alternatív nevüket még mindig használják. Így a p21 fehérje WAF1 (vad típusú p53 aktivált 1. fragmentum), CIP1 (CDK2 interacting protein 1), SDI1 (öregedésből származó inhibitor 1) és mda-6 (melanoma differenciálódáshoz társuló gén) néven is ismert. A p27 és p57 szinonimái a KIP1 (kinázt gátló fehérjék 1) és a KIP2 (kinázt gátló fehérjék 2). Mindezek a fehérjék széles specifikus hatást fejtenek ki, és gátolhatnak különféle CDK .
Ezzel szemben az INK4 inhibitorok csoportja specifikusabb. Négy fehérjét tartalmaz: p15INK4B , p16INK4A , p18INK4CÉs p19INK4D. Az INK4 család inhibitorai a fázis alatt működnek G1 sejtciklus, gátolja az aktivitást CDK4 kináz, azonban a második az INK4A gén fehérje terméke - p19ARF, interakcióba lép vele szabályozó faktor MDM2 fehérje p53és inaktiválja a faktort. Ez fokozott stabilitással jár együtt p53 fehérjeés állj meg
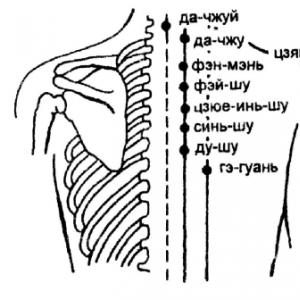
Sejtciklus: a G1-ből az S-fázisba való átmenet szabályozása
A sejtciklus kezdete előtt p27 fehérje, mivel nagy koncentrációban van, megakadályozza az aktiválódást protein kináz CDK4 vagy CDK6 ciklinek D1 , D2 vagy D3. Ilyen körülmények között a sejt bent marad fázis G0 vagy korai G1 fázis mielőtt megkapná a mitogén ingert. Megfelelő stimuláció után a p27-inhibitor koncentrációja csökken a ciklin D intracelluláris tartalmának növekedése miatt. Ezt a CDK aktiválódása és végső soron foszforiláció kíséri. pRb fehérje, a kapcsolódó kiadása transzkripciós faktor E2Fés a megfelelő gének transzkripciójának aktiválása.
A sejtciklus G1 fázisának korai szakaszában a p27 fehérje koncentrációja még mindig meglehetősen magas. Ezért a sejtek mitogén stimulációjának megszűnése után e fehérje tartalma gyorsan visszaáll a kritikus szintre, és a sejtek további áthaladása a sejtcikluson a megfelelő G1 szakaszban blokkolva van. Ez a visszafordíthatóság mindaddig lehetséges, amíg a G1 fázis fejlődésében el nem ér egy bizonyos ún átmeneti pont, ami után a sejt elköteleződik az osztódás mellett, és a növekedési faktorok környezetből való eltávolítása nem jár együtt a sejtciklus gátlásával. Bár ettől kezdve a sejtek függetlenné válnak az osztódásra irányuló külső jelektől, megtartják a sejtciklus önszabályozásának képességét.
Az INK4 család CDK-gátlói (p15 , 16. o , 18. oÉs 19. o) kifejezetten interakcióba lép CDK4 kinázokÉs CDK6. A p15 és p16 fehérjéket tumorszuppresszorként azonosították, és szintézisük szabályozott pRb fehérje. Mind a négy fehérje blokkolja a CDK4 és CDK6 aktiválását, vagy gyengíti a ciklinekkel való kölcsönhatásukat, vagy kiszorítja őket a komplexből. Bár mind a p16, mind a p27 fehérjék képesek gátolni a CDK4 és CDK6 aktivitását, az előbbi nagyobb affinitást mutat ezekhez a protein kinázokhoz. Ha a p16 koncentrációja olyan szintre emelkedik, amelynél teljesen gátolja a CDK4/6 kinázok aktivitását, a p27 fehérje lesz a fő inhibitor. CDK2 kináz .
A sejtciklus korai szakaszában az egészséges sejtek képesek felismerni és reagálni a DNS-károsodásra azáltal, hogy leállítják a sejtciklus progresszióját a G1 fázisban, amíg a károsodást ki nem javítják. Például az ultraibolya fény vagy ionizáló sugárzás által okozott DNS-károsodásra válaszul, p53 fehérje transzkripciót indukál p21 fehérje gén. Az intracelluláris koncentrációjának növelése blokkolja a CDK2 aktiválódását ciklinek E vagy A. Ez leállítja a sejteket a sejtciklus késői G1 fázisában vagy korai S fázisában. Ekkor maga a sejt határozza meg jövőbeli sorsát - ha a károsodást nem lehet megszüntetni, akkor belemegy apoptózis .
Két eltérő irányítású szabályozási rendszer létezik G1/S- átmenet: pozitív és negatív.
A rendszer pozitívan szabályozza az S-fázisba való belépést, tartalmaz egy heterodimert E2F-1/DP-1és aktiválja azt ciklin kináz komplexek .
Egy másik rendszer gátolja az S-fázisba való belépést. Ezt a tumorszuppresszorok képviselik p53És pRB, amelyek elnyomják az E2F-1/DP-1 heterodimerek aktivitását.
A normális sejtproliferáció e rendszerek közötti pontos egyensúlytól függ. Ezeknek a rendszereknek a kapcsolata megváltozhat, ami a sejtproliferáció sebességének változásához vezethet.
Sejtciklus: a G2 fázisból az M fázisba való átmenet szabályozása
A sejt DNS-károsodásra adott válasza már korábban is előfordulhat mitózis. Akkor p53 fehérje inhibitor szintézist indukál p21, ami megakadályozza az aktiválást
CDK1 kináz ciklin Bés késlelteti a sejtciklus további fejlődését. A sejt mitózison való áthaladása szigorúan ellenőrzött - a következő szakaszok nem kezdődnek el az előzőek teljes befejezése nélkül. Az inhibitorok egy részét élesztőben azonosították, de állati homológjaik ismeretlenek. Például leírt élesztőfehérjék BUB1 (a bimbózást nem gátolja a benomil)És MAD2 (mitotikus leállás hiányos), amelyek szabályozzák a kondenzált kromoszómák kapcsolódását a mitotikus orsóhoz a mitózis metafázisa. Mielőtt ezeknek a komplexeknek a megfelelő összeállítása befejeződik, a MAD2 fehérje komplexet képez a protein kináz CDC20és inaktiválja azt. A CDC20 aktiválás után foszforilálja a fehérjéket, és ennek eredményeként blokkolja azokat a funkcióit, amelyek megakadályozzák a két homológ kromatid eltérését citokinézis .
Következtetés
Élesztő- és emlőssejtvonalak hőmérsékletfüggő mutánsaival végzett kísérletek kimutatták, hogy a mitózis előfordulását bizonyos gének aktiválása, valamint specifikus RNS és fehérje szintézise határozza meg. Néha a mitózist csak nukleáris osztódásnak (kariokinézisnek) tekintik, amelyet nem mindig kísér citotómia - két rész kialakulása. sejteket.
Így a mitózis következtében egy sejt kettővé alakul, amelyek mindegyikében az adott szervezettípusra jellemző kromoszómák száma és alakja, következésképpen állandó mennyiségű DNS-e van.
A mitózis biológiai jelentősége abban rejlik, hogy biztosítja a kromoszómák számának állandóságát a szervezet minden sejtjében. A mitózis folyamata során az anyasejt kromoszómáinak DNS-e szigorúan egyenlően oszlik el a belőle származó két leánysejt között. A mitózis eredményeként a nemi sejtek kivételével a test minden sejtje ugyanazt a genetikai információt kapja. Az ilyen sejteket szomatikusnak nevezik (a görög "soma" - test). ciklus). Sejtes ciklus- Ez az időszak... Mitotikus ciklus magában foglalja a mitózist, valamint a pihenőidőt (G0), a posztmitózist ( G1), szintetikus (S) és premitotikus ( G2... . posztmitózisos időszak ( G1). Fázis G1- ez a fő...
A sejtek létezése időben és térben. Sejtes ciklusés annak szabályozása
Teszt >> BiológiaMegosztás vagy halál. Mitotikusés az élet ciklus egybeesik a gyakori felosztásban... (30-40% sejtes ciklus) erősödik. Után G1 fázisok S-vel kezdődik fázis. Pontosan az történik... . A szaporodás utáni helyreállítás történik G2 fázis. BAN BEN G2 fázis(10-20%) szintézis megy végbe...
Élet ( sejtes) ciklus
Jelentés >> BiológiaLétfontosságúnak, ill sejtes ciklus. Az újonnan előkerült sejt... mitotikus. Az interfázis viszont három időszakot foglal magában: preszintetikus - G1, szintetikus - S és posztszintetikus - G2. preszintetikus ( G1...ez fázisok körülbelül 4 óra.
A modern genetika fejlődéstörténete és főbb eredményei
Absztrakt >> Orvostudomány, egészségügyA sejt időbeli szerveződése. SejtesÉs mitotikus ciklusok. Sejtes ciklus– ez az időszak... Preszintetikus (posztmitotikus) G1– időtartam tól... . c) Posztszintetikus időszak G2– időtartama kisebb, ... 4-re osztva fázisok: profázis, metafázis, ...
Sejtciklus a sejt életének egyik szakasza az egyik osztódástól a másikig. Interfázisokból és felosztási periódusokból áll. A sejtciklus időtartama különböző szervezetekben változik (baktériumok esetében - 20-30 perc, eukarióta sejtek esetében - 10-80 óra).
Interfázis
Interfázis (a lat. inter- között, fázisok– felbukkanás) a sejtosztódások közötti vagy az osztódástól a haláláig eltelt időszak. A sejtosztódástól a haláláig eltelt időszak egy többsejtű szervezet azon sejtjeire jellemző, amelyek az osztódás után elveszítették erre való képességüket (eritrociták, idegsejtek stb.). Az interfázis a sejtciklus körülbelül 90%-át veszi fel.
Az interfázis tartalmazza:
1) preszintetikus időszak (G 1) – intenzív bioszintézis folyamatok indulnak be, a sejt növekszik és megnő. Ebben az időszakban maradnak meg a többsejtű szervezetek osztódási képességét elvesztett sejtjei a halálig;
2) szintetikus (S) – a DNS és a kromoszómák megduplázódnak (a sejt tetraploid lesz), a centriolok, ha vannak, megduplázódnak;
3) posztszintetikus (G 2) – a sejtben alapvetően leállnak a szintézis folyamatok, a sejt felkészül az osztódásra.
Megtörténik a sejtosztódás közvetlen(amitózis) és közvetett(mitózis, meiózis).
Amitózis
Amitózis – közvetlen sejtosztódás, amelyben nem alakul ki osztódási apparátus. A mag a gyűrűs szűkület miatt osztódik. A genetikai információnak nincs egységes eloszlása. A természetben az emlősök csillós sejtjeinek és méhlepényeinek makronukleuszait (nagy magvakat) amitózis osztja szét. A rákos sejtek amitózissal osztódhatnak.
A közvetett osztódás a hasadási apparátus kialakulásához kapcsolódik. Az osztókészülék olyan komponenseket tartalmaz, amelyek biztosítják a kromoszómák egyenletes eloszlását a sejtek között (osztódási orsó, centromerek és, ha vannak, centriolák). A sejtosztódás nukleáris osztódásra osztható ( mitózis) és a citoplazmatikus osztódás ( citokinézis). Ez utóbbi az atommaghasadás vége felé kezdődik. A természetben a leggyakoribb a mitózis és a meiózis. Alkalmanként előfordul endomitózis- közvetett hasadás, amely a magban a héj elpusztítása nélkül megy végbe.
Mitózis
Mitózis egy közvetett sejtosztódás, amelyben az anyasejtből két azonos genetikai információkészlettel rendelkező leánysejt képződik.
Mitózis fázisai:
1) próféta – kromatin tömörödés (kondenzáció) következik be, a kromatidák spiráloznak és megrövidülnek (fénymikroszkópban láthatóvá válnak), a nukleolusok és a magmembrán eltűnik, orsó keletkezik, fonalai a kromoszómák centromereihez kapcsolódnak, centriolok osztódnak és szétválnak a pólusok felé a sejt;
2) metafázis – a kromoszómák maximálisan spiralizáltak és az egyenlítő mentén helyezkednek el (az egyenlítői lemezben), a homológ kromoszómák a közelben helyezkednek el;
3) anafázis – az orsószálak egyszerre húzódnak össze, és a kromoszómákat a pólusokig nyújtják (a kromoszómák monokromatidá válnak), a mitózis legrövidebb fázisa;
4) telofázis – a kromoszómák despirálnak, sejtmagok és magmembrán képződnek, megindul a citoplazma osztódása.
A mitózis elsősorban a szomatikus sejtekre jellemző. A mitózis állandó számú kromoszómát tart fenn. Segíti a sejtek számának növelését, ezért megfigyelhető a növekedés, a regeneráció és a vegetatív szaporodás során.
Meiosis
Meiosis (görögből meiózis- redukció) egy közvetett redukciós sejtosztódás, amelyben az anyasejtből négy, nem azonos genetikai információval rendelkező leánysejt jön létre.
Két felosztása van: meiosis I. és meiosis II. Az I. interfázis hasonló a mitózis előtti interfázishoz. Az interfázis szintetikus utáni periódusában a fehérjeszintézis folyamatai nem állnak le, és az első osztódás profázisában folytatódnak.
Meiosis I:
– Prófázis I – a kromoszómák spiráloznak, a sejtmag és a magburok eltűnik, orsó képződik, a homológ kromoszómák közelebb kerülnek egymáshoz és a testvérkromatidák mentén összetapadnak (mint villám a kastélyban) – előfordul konjugáció, így kialakul tetradák, vagy bivalensek kromoszómák keresztezése és metszetcsere jön létre - átkelés, akkor a homológ kromoszómák taszítják egymást, de kapcsolódva maradnak azokon a területeken, ahol az átkelés megtörtént; a szintézis folyamatok befejeződnek;
– metafázis I – a kromoszómák az Egyenlítő mentén helyezkednek el, homológok – a bikromatid kromoszómák egymással szemben helyezkednek el az Egyenlítő mindkét oldalán;
– anafázis I – az orsó filamentumai egyszerre húzódnak össze és nyúlnak egy homológ bikromatid kromoszóma mentén a pólusok felé;
– telofázis I (ha van) - a kromoszómák despirálnak, sejtmag és nukleáris membrán képződik, a citoplazma eloszlik (a képződő sejtek haploidok).
Interfázis II(ha van): DNS-duplikáció nem fordul elő.
Meiosis II:
– prófázis II – a kromoszómák sűrűsödnek, a sejtmag és a magmembrán eltűnik, hasadási orsó alakul ki;
– metafázis II – a kromoszómák az Egyenlítő mentén helyezkednek el;
– anafázis II – a kromoszómák az orsószálak egyidejű összehúzódásával a pólusokhoz térnek el;
– telofázis II – a kromoszómák despirálnak, sejtmag és magmembrán keletkezik, a citoplazma osztódik.
A meiózis a csírasejtek kialakulása előtt következik be. Lehetővé teszi a csírasejtek fúzióját a faj állandó számú kromoszómájának fenntartásához (kariotípus). Kombinatív variálhatóságot biztosít.