Mi a sejtfal része. Sejtfal
A sejtfal kemény, félig áteresztő védőréteg egyes sejttípusokban. Ez a külső borítás megtalálható a legtöbb növényi sejtben, gombában, baktériumban, algában és néhány archaeában. Az állatoknak azonban nincs sejtfaluk. Számos fontos funkciót lát el, beleértve a védelmet és a szerkezeti támogatást.
A sejtfal szerkezetének jellemzői a szervezet típusától függenek. Például a növényekben általában erős szénhidrát-polimer cellulóz rostokból áll, amely a pamut és a fa fő összetevője, és a papírgyártásban is használják.
A növényi sejtfal szerkezete
A növények sejtfala többrétegű, és három részből áll: a külső rétegből vagy a középső rétegből, az elsődleges és a másodlagos sejtfalból. Bár minden növényi sejtnek van középső rétege és elsődleges sejtfala, nem minden növényi sejtnek van másodlagos sejtfala.
A középső réteg a sejtfal külső rétege, amely poliszacharidokat, úgynevezett pektineket tartalmaz. A pektinek segítik a sejtek adhézióját azáltal, hogy egymáshoz kötik a szomszédos sejtek falát.
Az elsődleges sejtfal a középső lamina és a plazmamembrán között kialakuló réteg a növekvő növényi sejtekben. Főleg cellulóz mikrofibrillumokból áll, amelyeket hemicellulóz rostok és pektin poliszacharidok gélmátrixa tartalmaz. Az elsődleges sejtfal biztosítja a sejtnövekedéshez szükséges erőt és rugalmasságot.
A másodlagos sejtfal egy réteg, amely egyes növényi sejtekben az elsődleges sejtfal és a plazmamembrán között képződik. Amikor az elsődleges sejtfal abbahagyja az osztódást és növekedést, megvastagodhat, és másodlagos sejtfalat képezhet. Ez a tartós réteg megerősíti és alátámasztja a ketrecet. Egyes másodlagos sejtfalakban a cellulóz és a hemicellulóz mellett lignin is található, amely erősíti ezeket, és biztosítja a növények érszövetének sejtjeinek áteresztőképességét.
A sejtfal funkciói
A sejtfal fő funkciója, hogy állványt képezzen a sejt számára, és megakadályozza annak tágulását. A cellulózrostok, a szerkezeti fehérjék és más poliszacharidok formát adnak a sejteknek és támaszt nyújtanak. A sejtfal további funkciói a következők:
- Támogatás - mechanikai szilárdság és szerkezet biztosítása, valamint a sejtnövekedés irányának szabályozása.
- Ellenáll a turgornyomásnak - a sejt (protoplaszt) tartalmának a falára ható erejének. Ez a nyomás segít a növénynek merev és felálló maradni, de sejtpusztulást is okozhat.
- Növekedési szabályozás – Jeleket küld a sejteknek, hogy belépjenek, osztódjanak és növekedjenek.
- Diffúziós szabályozás – A sejtfal porózus szerkezete lehetővé teszi bizonyos esszenciális anyagok, köztük a fehérjék bejutását a sejtbe, megakadályozva mások bejutását.
- Kommunikáció – a sejtek kölcsönhatásba lépnek egymással a plazmodezmákon keresztül (a növényi sejtfalak közötti pórusok vagy csatornák, amelyek lehetővé teszik a molekulák és kommunikációs jelek átjutását az egyes növényi sejtek között).
- Védelem - megvédi a sejteket a vírusoktól és más veszélyes anyagoktól vagy mikroorganizmusoktól, valamint segít megelőzni a vízveszteséget.
- Tárolás - szénhidrátokat tárol, amelyeket a növények növekedéséhez használnak, különösen a magvakban.
A földi élet kialakulásának hajnalán minden sejtformát baktériumok képviseltek. A test felszínén keresztül szívták magukba az elsődleges óceánban oldott szerves anyagokat.
Idővel egyes baktériumok alkalmazkodtak ahhoz, hogy szervetlen anyagokból szerves anyagokat állítsanak elő. Ehhez a napfény energiáját használták fel. Létrejött az első ökológiai rendszer, amelyben ezek az organizmusok termelők voltak. Ennek eredményeként oxigén jelent meg a Föld légkörében, amelyet ezek az élőlények bocsátanak ki. Segítségével ugyanabból a táplálékból sokkal több energiát lehet nyerni, a plusz energiával pedig a test felépítését lehet bonyolítani: a testet részekre osztani.
Az élet egyik fontos vívmánya a sejtmag és a citoplazma szétválása. A mag örökletes információkat tartalmaz. A mag körül található speciális membrán lehetővé tette a véletlen sérülések elleni védelmet. Szükség esetén a citoplazma parancsokat kap a sejtmagtól, amelyek irányítják a sejt létfontosságú tevékenységét és fejlődését.
Azok az élőlények, amelyekben a sejtmag elválik a citoplazmától, a nukleáris szuperbirodalomot alkották (ide tartoznak a növények, gombák, állatok).
Így a sejt - a növények és állatok szerveződésének alapja - a biológiai evolúció során keletkezett és fejlődött.
Még szabad szemmel, de még jobb nagyító alatt is láthatja, hogy az érett görögdinnye pépje nagyon apró szemekből, vagy szemekből áll. Ezek a sejtek - a legkisebb „téglák”, amelyek az összes élő szervezet testét alkotják, beleértve a növényieket is.
A növény életét egyetlen egészet létrehozó sejtjei együttes tevékenysége hajtja végre. A növényi részek többsejtűségével funkcióik fiziológiai differenciálódása, a különféle sejtek specializálódása a növényi testben való elhelyezkedésük függvényében következik be.
A növényi sejt abban különbözik az állati sejttől, hogy sűrű héja van, amely minden oldalról befedi a belső tartalmat. A sejt nem lapos (ahogy általában ábrázolják), valószínűleg úgy néz ki, mint egy nagyon kicsi, nyálkahártya-tartalommal teli hólyag.
A növényi sejt szerkezete és működése
Tekintsük a sejtet egy szervezet szerkezeti és funkcionális egységének. Kívül a sejtet sűrű sejtfal borítja, amelyben vékonyabb területek - pórusok - vannak. Alatta van egy nagyon vékony film - egy membrán, amely lefedi a sejt tartalmát - a citoplazmát. A citoplazmában üregek vannak - sejtnedvvel töltött vakuolák. A sejt közepén vagy a sejtfal közelében egy sűrű test található - egy mag egy maggal. A sejtmagot magburok választja el a citoplazmától. A kis testek - plasztiszok - a citoplazmában eloszlanak.
Növényi sejt szerkezete
A növényi sejtszervecskék felépítése és működése
| Organoid | Rajz | Leírás | Funkció | Sajátosságok |
Sejtfal vagy plazmamembrán | Színtelen, átlátszó és nagyon tartós | Beengedi az anyagokat a sejtbe és onnan ki. | A sejtmembrán félig áteresztő |
|
Citoplazma | Vastag viszkózus anyag | A sejt összes többi része benne található. | Állandó mozgásban van |
|
A sejtmag (a sejt fontos része) | Lekerekített vagy ovális | Biztosítja az örökletes tulajdonságok átadását a leánysejteknek az osztódás során | A sejt központi része |
|
Gömb alakú vagy szabálytalan alakú | Részt vesz a fehérjeszintézisben | |||
 | A citoplazmától membránnal elválasztott rezervoár. Sejtlevet tartalmaz | Tartalék tápanyagok és salakanyagok halmozódnak fel, amelyek a sejt számára feleslegesek. | Ahogy a sejt növekszik, a kis vakuolák egyetlen nagy (központi) vakuólummá egyesülnek |
|
Plasztidok | Kloroplasztok | Használja ki a nap fényenergiáját, és alkosson szerves anyagokat szervetlen anyagokból | A citoplazmától kettős membránnal határolt korongok alakja |
|
Kromoplasztok | A karotinoidok felhalmozódása eredményeként képződik | Sárga, narancs vagy barna |
||
 | Leukoplasztok | Színtelen plasztidok | ||
Nukleáris héj | Két membránból áll (külső és belső), pórusokkal | Elválasztja a sejtmagot a citoplazmától | Lehetővé teszi a sejtmag és a citoplazma közötti cserét |
A sejt élő része egy membránnal határolt, rendezett, strukturált biopolimerekből és belső membránstruktúrákból álló rendszer, amely az egész rendszer egészét fenntartó és újratermelő anyagcsere- és energiafolyamatok halmazában vesz részt.
Fontos jellemzője, hogy a sejtben nincsenek szabad végű nyitott membránok. A sejtmembránok mindig behatárolják az üregeket vagy területeket, minden oldalról lezárva azokat.
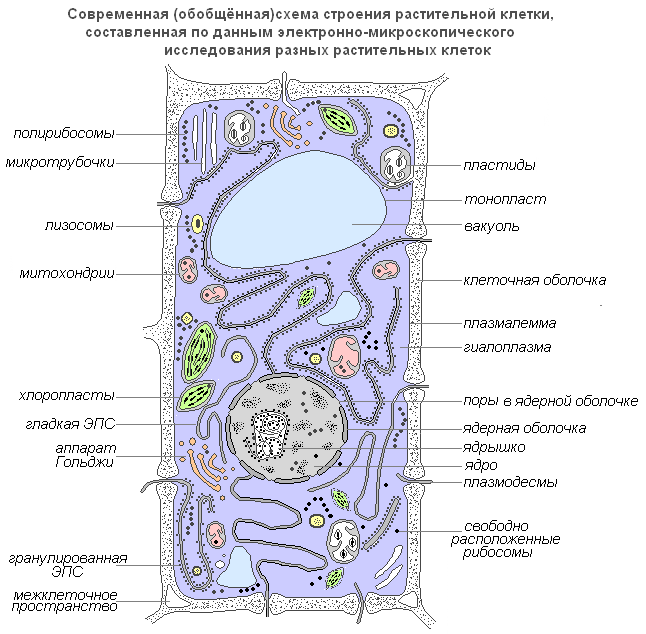
Egy növényi sejt modern általánosított diagramja
Plasmalemma(külső sejtmembrán) - 7,5 nm vastag ultramikroszkópos film, amely fehérjékből, foszfolipidekből és vízből áll. Ez egy nagyon rugalmas fólia, amely jól nedvesíthető vízzel, és gyorsan visszaállítja integritását sérülés után. Univerzális szerkezetű, azaz minden biológiai membránra jellemző. A növényi sejtekben a sejtmembránon kívül van egy erős sejtfal, amely külső támaszt nyújt és megtartja a sejt alakját. Rostból (cellulózból), vízben oldhatatlan poliszacharidból áll.
Plasmodesmata A növényi sejtek olyan szubmikroszkópos tubulusok, amelyek áthatolnak a membránokon, és plazmamembránnal vannak bélelve, amely így megszakítás nélkül átjut egyik sejtből a másikba. Segítségükkel megtörténik a szerves tápanyagokat tartalmazó oldatok sejtközi keringése. Biopotenciálok és egyéb információk átvitelére is használják őket.
Pórusúgynevezett lyukak a másodlagos membránban, ahol a sejteket csak az elsődleges membrán és a medián lemez választja el. A primer membrán és a szomszédos sejtek szomszédos pórusait elválasztó median lamina területeit pórusmembránnak vagy póruszáró filmnek nevezzük. A pórusok zárófilmje behatol a plazmodizálható tubulusokba, de átmenő lyuk általában nem képződik a pórusokban. A pórusok megkönnyítik a víz és az oldott anyagok szállítását sejtről sejtre. A szomszédos sejtek falában rendszerint egymás ellen pórusok képződnek.
Sejt membrán jól körülhatárolható, viszonylag vastag poliszacharid jellegű héja van. A növényi sejtmembrán a citoplazma aktivitásának terméke. A Golgi-készülék és az endoplazmatikus retikulum aktívan részt vesz a kialakulásában.

A sejtmembrán szerkezete
A citoplazma alapja a mátrixa, vagy hialoplazmája, egy összetett színtelen, optikailag átlátszó kolloid rendszer, amely képes reverzibilis átmenetekre szolból gélbe. A hialoplazma legfontosabb szerepe az összes sejtszerkezet egyetlen rendszerré egyesítése, és a köztük lévő kölcsönhatás biztosítása a sejtanyagcsere folyamataiban.
Hialoplazma(vagy a citoplazma mátrixa) alkotja a sejt belső környezetét. Vízből és különböző biopolimerekből (fehérjék, nukleinsavak, poliszacharidok, lipidek) áll, amelyek fő részét különböző kémiai és funkcionális specifitású fehérjék teszik ki. A hialoplazma aminosavakat, monoszacharidokat, nukleotidokat és más alacsony molekulatömegű anyagokat is tartalmaz.
A biopolimerek vízzel kolloid közeget képeznek, amely a körülményektől függően lehet sűrű (gél formájában) vagy folyékonyabb (szol formájában), mind a teljes citoplazmában, mind annak egyes területein. A hialoplazmában különböző organellumok és zárványok lokalizálódnak, és kölcsönhatásba lépnek egymással és a hialoplazma környezetével. Sőt, elhelyezkedésük leggyakrabban bizonyos típusú sejtekre jellemző. A bilipid membránon keresztül a hialoplazma kölcsönhatásba lép az extracelluláris környezettel. Következésképpen a hialoplazma dinamikus környezet, és fontos szerepet játszik az egyes organellumok működésében és általában a sejtek létfontosságú tevékenységében.
Citoplazmatikus képződmények - organellumok
Az organellumok (organellumok) a citoplazma szerkezeti alkotóelemei. Bizonyos alakúak és méretűek, a sejt kötelező citoplazmatikus struktúrái. Hiányuk vagy károsodásuk esetén a sejt rendszerint elveszíti továbbléti képességét. Sok organellum képes osztódásra és önszaporodásra. Méreteik olyan kicsik, hogy csak elektronmikroszkóppal láthatók.
Mag
A sejtmag a sejt leginkább látható és általában a legnagyobb organellumja. Először Robert Brown vizsgálta meg részletesen 1831-ben. A sejtmag biztosítja a sejt legfontosabb metabolikus és genetikai funkcióit. Meglehetősen változó alakú: lehet gömb alakú, ovális, karéjos, lencsés.
A sejtmag jelentős szerepet játszik a sejt életében. A sejt, amelyből a sejtmagot eltávolították, már nem választ ki membránt, abbahagyja a növekedést és az anyagok szintetizálását. Megszaporodnak benne a bomlás és a pusztulás termékei, aminek következtében gyorsan elpusztul. Új sejtmag képződése a citoplazmából nem következik be. Új magok csak a régi osztódásával vagy összezúzásával jönnek létre.
A mag belső tartalma a kariolimfa (maglé), amely kitölti a mag szerkezetei közötti teret. Egy vagy több sejtmagot, valamint jelentős számú DNS-molekulát tartalmaz, amelyek specifikus fehérjékhez - hisztonokhoz - kapcsolódnak.

A mag felépítése
Nucleolus
A sejtmag a citoplazmához hasonlóan főleg RNS-t és specifikus fehérjéket tartalmaz. Legfontosabb funkciója, hogy riboszómákat képez, amelyek a fehérjék szintézisét végzik a sejtben.
Golgi készülék
A Golgi-készülék egy organoid, amely általánosan elterjedt minden típusú eukarióta sejtben. Ez a lapos membrántasakok többszintű rendszere, amelyek a periféria mentén megvastagodnak és hólyagos folyamatokat képeznek. Leggyakrabban a mag közelében található.

Golgi készülék
A Golgi-készülék szükségszerűen tartalmaz egy kis hólyagok (vezikulák) rendszerét, amelyek a megvastagodott ciszternákból (korongokból) válnak le, és ennek a szerkezetnek a perifériáján helyezkednek el. Ezek a vezikulák specifikus szektorszemcsék intracelluláris transzportrendszerének szerepét töltik be, és celluláris lizoszómák forrásaként szolgálhatnak.
A Golgi-készülék funkciói az intracelluláris szintézis termékeinek, a bomlástermékeknek és a toxikus anyagoknak a buborékok segítségével történő sejten kívüli felhalmozódásában, elválasztásában és kiválasztásában is szerepelnek. A sejt szintetikus aktivitásának termékei, valamint az endoplazmatikus retikulum csatornáin keresztül a környezetből a sejtbe jutó különféle anyagok a Golgi-készülékbe kerülnek, ebben az organoidban halmozódnak fel, majd cseppek vagy szemcsék formájában. bejutnak a citoplazmába, és vagy maga a sejt használja fel őket, vagy kívülről ürülnek ki... A növényi sejtekben a Golgi-készülék a poliszacharidok szintéziséhez szükséges enzimeket és magát a poliszacharid anyagot tartalmazza, amelyet a sejtfal építésére használnak. Úgy gondolják, hogy részt vesz a vakuolák kialakulásában. A Golgi-készüléket Camillo Golgi olasz tudósról nevezték el, aki 1897-ben fedezte fel először.
Lizoszómák
A lizoszómák membránnal határolt kis vezikulák, amelyek fő funkciója az intracelluláris emésztés. A lizoszómális apparátus alkalmazása a növény magjának csírázása során történik (tartalék tápanyagok hidrolízise).

Lizoszóma szerkezete
Mikrotubulusok
A mikrotubulusok membrán, szupramolekuláris struktúrák, amelyek spirális vagy egyenes vonalú sorokban elhelyezkedő fehérjegömbökből állnak. A mikrotubulusok elsősorban mechanikai (motoros) funkciót látnak el, biztosítva a sejtszervecskék mobilitását és kontraktilitását. A citoplazmában találhatók, bizonyos formát adnak a sejtnek, és biztosítják az organellumok térbeli elrendezésének stabilitását. A mikrotubulusok elősegítik az organellumok mozgását olyan helyekre, amelyeket a sejt fiziológiai szükségletei határoznak meg. Ezen struktúrák jelentős része a plazmalemmában, a sejtmembrán közelében található, ahol részt vesznek a növényi sejtmembránok cellulóz mikrofibrillumainak kialakításában és orientációjában.

Mikrotubulus szerkezet
Vacuole
A vakuólum a növényi sejtek legfontosabb összetevője. Ez egyfajta üreg (tartály) a citoplazma tömegében, amelyet ásványi sók, aminosavak, szerves savak, pigmentek, szénhidrátok vizes oldatával töltenek meg, és egy vakuoláris membrán - tonoplaszt - választják el a citoplazmától.
A citoplazma csak a legfiatalabb növényi sejtekben tölti ki a teljes belső üreget. A sejt növekedésével a kezdetben folytonos citoplazmatömeg térbeli elrendeződése jelentősen megváltozik: sejtnedvvel teli kis vakuolák jelennek meg benne, és a teljes tömeg szivacsossá válik. A további sejtnövekedéssel az egyes vakuolák egyesülnek, a citoplazma rétegeit a perifériára tolják, aminek következtében a kialakult sejtben általában egy nagy vakuólum van, és a citoplazma az összes organellával a membrán közelében helyezkedik el.
A vakuolák vízben oldódó szerves és ásványi vegyületei határozzák meg az élő sejtek megfelelő ozmotikus tulajdonságait. Ez a bizonyos koncentrációjú oldat egyfajta ozmotikus pumpa a sejtbe történő szabályozott behatoláshoz, valamint víz, ionok és metabolitmolekulák felszabadulásához.
A féligáteresztő tulajdonságokkal jellemezhető citoplazmaréteggel és membránjaival kombinálva a vakuólum hatékony ozmotikus rendszert alkot. Az élő növényi sejtek olyan mutatóit, mint az ozmotikus potenciál, a szívóerő és a turgornyomás, ozmotikusan határozzák meg.

Vacuole szerkezet
Plasztidok
A plasztidák a legnagyobb (a sejtmag után) citoplazmaszervecskék, amelyek csak a növényi szervezetek sejtjeiben rejlenek. Nem csak a gombákban találhatók meg. A plasztidok fontos szerepet játszanak az anyagcserében. A citoplazmától kettős membránmembrán választja el őket, egyes típusaik jól fejlett és rendezett belső membránrendszerrel rendelkeznek. Minden plasztid azonos eredetű.
Kloroplasztok- a fotoautotróf szervezetek legelterjedtebb és funkcionálisan legfontosabb plasztidjai, amelyek olyan fotoszintetikus folyamatokat hajtanak végre, amelyek végső soron szerves anyagok képződéséhez és szabad oxigén felszabadulásához vezetnek. A magasabb rendű növények kloroplasztjai összetett belső szerkezettel rendelkeznek.
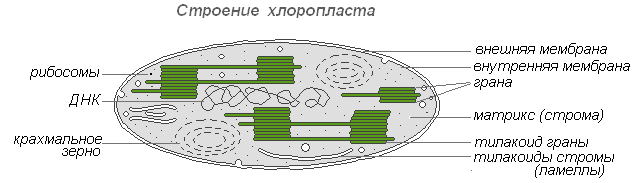
Kloroplaszt szerkezet
A kloroplasztiszok méretei a különböző növényekben nem azonosak, de átlagosan 4-6 mikron átmérőjűek. A kloroplasztok képesek mozogni a citoplazma mozgásának hatására. Ezenkívül a megvilágítás hatására az amőba típusú kloroplasztiszok aktív mozgása zajlik a fényforrás felé.
A klorofill a kloroplasztiszok fő anyaga. A klorofillnak köszönhetően a zöld növények képesek fényenergiát hasznosítani.
Leukoplasztok(színtelen plasztidok) egyértelműen megjelölt citoplazmatikus testek. Méretük valamivel kisebb, mint a kloroplasztiszok mérete. Egyre monotonabbak és formájuk, közelebb a gömb alakúhoz.

Leukoplaszt szerkezet
Az epidermisz sejtjeiben, gumókban, rizómákban található. Megvilágításkor nagyon gyorsan kloroplasztiszokká alakulnak, és ennek megfelelően megváltoznak a belső szerkezetük. A leukoplasztok enzimeket tartalmaznak, amelyek segítségével a fotoszintézis során képződő glükózfeleslegből keményítő szintetizálódik, amelynek nagy része a tárolószövetekben vagy szervekben (gumók, rizómák, magvak) keményítőszemcsék formájában rakódik le. Egyes növényekben a zsírok leukoplasztokban rakódnak le. A leukoplasztok tartalék funkciója esetenként kristályok vagy amorf zárványok formájában raktározó fehérjék képződésében nyilvánul meg.
Kromoplasztok a legtöbb esetben kloroplasztiszok származékai, esetenként leukoplasztok.

A kromoplaszt szerkezete
A csipkebogyó, paprika, paradicsom érése a pépsejtek kloro- vagy leukoplasztjainak karotinoidoplasztokká történő átalakulásával jár. Ez utóbbiak főleg sárga plasztid pigmenteket - karotinoidokat tartalmaznak, amelyek érett állapotban intenzíven szintetizálódnak bennük, színes lipidcseppeket, szilárd gömböcskéket vagy kristályokat képezve. Ebben az esetben a klorofill megsemmisül.
Mitokondriumok
A mitokondriumok olyan organellumok, amelyek a legtöbb növényi sejtre jellemzőek. Változtatható formájú pálcikák, szemcsék, cérnák. 1894-ben fedezte fel R. Altman fénymikroszkóp segítségével, a belső szerkezetet később elektronmikroszkóppal tanulmányozták.

A mitokondrium szerkezete
A mitokondriumok két membránból állnak. A külső membrán sima, a belső különböző formájú kinövéseket képez - tubulusokat a növényi sejtekben. A mitokondriumok belsejében lévő teret félig folyékony tartalom (mátrix) tölti ki, amely enzimeket, fehérjéket, lipideket, kalcium- és magnéziumsókat, vitaminokat, valamint RNS-t, DNS-t és riboszómákat tartalmaz. A mitokondriumok enzimatikus komplexe felgyorsítja az ATP képződését eredményező biokémiai reakciók összetett és egymással összefüggő mechanizmusát. Ezekben az organellumokban a sejteket energiával látják el - a tápanyagok kémiai kötéseinek energiáját nagy energiájú ATP-kötésekké alakítják a sejtlégzés folyamatában. A mitokondriumokban zajlik le a szénhidrátok, zsírsavak, aminosavak enzimatikus lebontása az energia felszabadulásával és az azt követő ATP energiává történő átalakulásával. A felhalmozott energiát növekedési folyamatokra, új szintézisekre stb. fordítják. A mitokondriumok hasadással szaporodnak és körülbelül 10 napig élnek, majd elpusztulnak.
Endoplazmatikus retikulum
Az endoplazmatikus retikulum csatornák, tubulusok, vezikulák, ciszternák hálózata, amelyek a citoplazmában helyezkednek el. K. Porter angol tudós fedezte fel 1945-ben, ultramikroszkópos szerkezetű membránrendszer.

Az endoplazmatikus retikulum szerkezete
A teljes hálózat egyetlen egésszé épül fel a nukleáris burok külső sejtmembránjával. Tegyen különbséget a sima és durva EPS között, amely riboszómákat tartalmaz. A zsír- és szénhidrát-anyagcserében részt vevő enzimrendszerek a sima EPS membránjain helyezkednek el. Ez a fajta membrán dominál a tárolóanyagokban (fehérjék, szénhidrátok, olajok) gazdag magsejtekben, a szemcsés EPS membránjához riboszómák kötődnek, és a fehérje molekula szintézise során a riboszómákkal rendelkező polipeptidlánc az EPS csatornába merül. Az endoplazmatikus retikulum funkciói igen változatosak: anyagok szállítása a sejten belül és a szomszédos sejtek között egyaránt; a sejt külön szakaszokra osztása, amelyekben különböző élettani folyamatok és kémiai reakciók zajlanak egyszerre.
Riboszómák
A riboszómák nem membrán sejtszervecskék. Minden riboszóma két, nem azonos méretű részecskebõl áll, és két részre osztható, amelyek továbbra is megõrzik fehérjeszintetizáló képességüket, miután egy teljes riboszómává egyesülnek.

Riboszóma szerkezet
A riboszómák a sejtmagban szintetizálódnak, majd elhagyják azt, átjutnak a citoplazmába, ahol az endoplazmatikus retikulum membránjainak külső felületéhez kapcsolódnak, vagy szabadon helyezkednek el. A szintetizált fehérje típusától függően a riboszómák önállóan működhetnek, vagy komplexekké - poliriboszómákká - egyesülhetnek.
Ellentétben az állatokkal és sok protozoonnal, növényekben, baktériumokban és gombákban szinte minden sejtnek van fala, amely a citoplazma membránon kívül fekszik, és megnövekedett szilárdsággal rendelkezik. Ennek a szerkezetnek a fő funkciója a támogatás és a védelem.
A sejtfalak (vagy sejtfalak) maguk a sejtek által szintetizált anyagokból épülnek fel. Kémiai összetételük különböző növényekben, gombákban és prokariótákban. Ezenkívül még ugyanabban az üzemben is a falak összetétele némileg eltérő a különböző sejteknél.
A növények sejtfala főleg cellulózból áll. A cellulóz egy poliszacharid, amelynek monomerje glükóz.
A bakteriális sejtfalak alapja a murein (peptidoglikánokra utal). Gram-pozitív baktériumokban a membrán különféle savakat tartalmaz, és maga a membrán szorosan kapcsolódik a citoplazmatikus membránhoz. Gram-negatív baktériumokban a membrán vékonyabb, és nem csatlakozik a membránhoz. A membrán és a burok között periplazmatikus tér képződik. Kívül a gram-negatív prokarióták sejtmembránját lipopoliszacharidból álló külső membrán veszi körül.
A gombákban a sejtfalak fő anyaga kitin, nem cellulóz.
A növényi sejtfal összetétele
A növényekben a leánysejtek fala már a szülő osztódása során kialakul. Ezt követően elsődlegesnek nevezik. Sok sejt később másodlagos membránt fejleszt ki.
Az elsődleges sejtmembrán abból áll cellulóz mikrofibrillumok más poliszacharidok mátrixába merítve. A cellulózszálak megkülönböztető jellemzője az erősségük. A cellulózmolekula egy hosszú poliszacharid lánc. Az egyes molekulák hidrogénkötéssel kötődnek egymáshoz egy mikroszálnak nevezett kötegben. Ezek a rostok alkotják a sejtfal vázát.
A sejtfal mátrixa poliszacharidokból áll pektinek és hemicellulózok, valamint számos más anyag (például fehérjék). A pektin anyagok a savas poliszacharidok egy csoportja, molekuláik nem csak lineárisak, hanem elágazóak is lehetnek. A hemicellulózok szintén a poliszacharidok vegyes csoportját alkotják. Lineáris molekuláik rövidebbek, mint a cellulóz.
A szomszédos növényi sejtek héja összekapcsolódik median lamina, amely magnézium- és kalcium-pektátokból áll, amelyekre a ragadósság jellemző.
A növények falának összetétele vizet tartalmaz (a tömeg több mint felét teszi ki), ami a poliszacharidok számos fizikai és kémiai tulajdonságát okozza.
Az üzem merev váza sok helyen tele van csatornákkal ( plazmodezma), amelyen keresztül az egyik sejt citoplazmája kapcsolódik a szomszédos sejtek citoplazmájához.
A levél mezofil sejtjeinek (és néhány másnak is) egész életük során csak elsődleges faluk van. A sejtek többségében további cellulózrétegekből álló másodlagos fal rakódik le az elsődleges membrán belső oldalán. Általában ekkor a sejt már differenciálódott és nem növekszik (kivétel például a kollenchima sejtek).
A másodlagos megvastagodás minden különálló rétegében a cellulóz mikrofibrillák azonos szögben (egymással párhuzamosan) helyezkednek el. A különböző rétegek azonban eltérő szöggel rendelkeznek, ami nagyobb szilárdságot biztosít.
A növényi sejtek egy része lignifikál (a xilem légcsőelemei, szklerenchima stb.). Ez a folyamat a fal intenzív lignifikációján alapul (kis mennyiségű lignin minden membránban jelen van). A lignin nem poliszacharid, hanem összetett polimer anyag. A ligninlerakódások különböző formájúak lehetnek (folyamatos, gyűrűs, spirális, hálós). Egyben tartja a cellulózt, nem engedi elmozdulni. A lignin nemcsak erőt ad, hanem további védelmet is nyújt a káros fizikai és kémiai tényezők ellen.
A sejtfal funkciói
A különböző sejtek héja együttesen biztosítja az egész növénynek és egyes részeinek mechanikai szilárdságát és támasztását. A sejtfalnak ez a funkciója hasonló az állatok csontvázának egyik funkciójához. Azonban nem ő az egyetlen.
A falak merevsége megakadályozza a sejtek megnyúlását és törését. Ennek eredményeként a fizikai törvények szerint a víz ozmózis útján juthat be a sejtekbe. A lágyszárú növények esetében a sejtek turgocentrikussága jelenti az egyetlen támaszt.
A cellulóz mikrofibrillumok korlátozzák a sejtek növekedését és meghatározzák azok alakját. Ha mikrofibrillumok gyűrűzik a sejtben, akkor az megnő (a rostok irányába).
Kialakulnak a kapcsolódó sejtfalak apoplast amelyen keresztül víz és ásványi anyagok mozognak. A plazmodezmák a különböző sejtek tartalmát egyetlen rendszerbe kötik - szimplaszt.
A xylem, tracheidák, szitacsövek edényeinek fala szállító funkciót lát el.
Az epidermális sejtek külső sejtfalait viasz (kutikula) borítja. Egyrészt megakadályozza a víz elpárolgását, másrészt a káros mikroorganizmusok bejutását.
Egyes növényekben, bizonyos sejtekben a membránok módosulnak, és a tápanyagok tárolására szolgálnak.
A sejtfal (sejtfal) a növényi sejt jellegzetes tulajdonsága, amely megkülönbözteti az állati sejttől. A sejtfal sajátos formát ad a sejtnek. A speciális táptalajokon tenyésztett növényi sejtek, amelyekről enzimatikusan eltávolítják a falat, mindig gömb alakúak. A sejtfal erőt ad a sejtnek és védi a protoplasztot, kiegyenlíti a turgornyomást és így megakadályozza a plazmalemma felszakadását. A sejtfalak összessége alkotja a belső vázat, amely megtámasztja a növény testét és mechanikai szilárdságot ad.
A sejtfal színtelen és átlátszó, könnyen átereszti a napfényt. Általában a falak vízzel telítettek. A víz és a benne oldott kis molekulatömegű vegyületek szállítása (transzport az apoplaszt mentén) a sejtfalrendszer mentén történik.
A sejtfal főként poliszacharidokból áll, amelyek vázanyagra és mátrixanyagra oszthatók.
A növényi sejtfal vázanyaga a cellulóz (rost), amely béta-1,4-D-glükán. A bioszférában a legnagyobb mennyiségben előforduló szerves anyag. A cellulózmolekulák nagyon hosszú, el nem ágazó láncok, egymással párhuzamosan, több tízes csoportokban helyezkednek el, és számos hidrogénkötés tartja össze őket. Ennek eredményeként mikrofibrillák képződnek, amelyek a fal szerkezeti vázát hozzák létre és meghatározzák annak szilárdságát. A cellulóz mikrofibrillumai csak elektronmikroszkópban láthatók, átmérőjük 10-30 nm, hossza eléri a több mikront.
A cellulóz oldhatatlan és nem duzzad vízben. Kémiailag nagyon inert, szerves oldószerekben, tömény lúgokban és híg savakban nem oldódik. A cellulóz mikrofibrillák rugalmasak és nagyon szakadásállóak (hasonlóan az acélhoz). Ezek a tulajdonságok határozzák meg a cellulóz és termékei széles körű alkalmazását. A világ szinte teljes egészében cellulózból álló pamutszál termelése 1,5 * 107 tonna évente. A cellulózból füstmentes port, acetát selymet és viszkózt, celofánt és papírt nyernek. A cellulózra adott kvalitatív reakciót klór-cink-jód reagenssel hajtják végre, a cellulóz sejtfala kékeslila színű.
A gombákban a sejtfal vázanyaga a kitin, egy glükózamin-maradékokból felépülő poliszacharid. A kitin még a cellulóznál is tartósabb.
A mikrofibrillákat amorf mátrixba, általában vízzel telített műanyag gélbe merítik. A mátrix poliszacharidok összetett keveréke, amelynek molekulái több különböző cukor maradékaiból állnak, és rövidebb és elágazó láncúak, mint a cellulózé. A mátrix poliszacharidok meghatározzák a sejtfal olyan tulajdonságait, mint az erős duzzadás, a víz és a benne oldott kis molekulatömegű vegyületek nagy áteresztőképessége, valamint a kationcserélő tulajdonságok. A mátrix poliszacharidokat két csoportra osztják - pektin anyagokra és hemicellulózokra.
A pektin anyagok erősen megduzzadnak vagy vízben oldódnak. Lúgok és savak könnyen tönkreteszik őket. A pektin anyagok legegyszerűbb képviselői a vízben oldódó pektinsavak - az alfa-D-galakturonsav polimerizációjának termékei (100 egységig), amelyek 1,4-kötésekkel kapcsolódnak lineáris láncokba (alfa-1,4-D-galakturonán). ). A pektinsavak (pektinek) az alfa-D-galakturonsav nagyobb molekulatömegű (100-200 egység) polimer vegyületei, amelyekben a karboxilcsoportok részben metiláltak. A pektátok és pektinátok a pektin- és pektinsavak kalcium- és magnéziumsói. A pektinsavak, pektátok és pektinátok cukrok és szerves savak jelenlétében vízben oldódnak, így sűrű géleket képeznek.
A növények sejtfala főként protopektineket tartalmaz - a metoxilezett poligalakturonsav nagy molekulatömegű polimerjei arabinánokkal és galaktánokkal, a kétszikű növényekben a galakturonánláncok kis mennyiségű ramnózt tartalmaznak. A protopektinek vízben oldhatatlanok.
A hemicellulózok semleges cukrok maradványaiból felépülő elágazó láncok, gyakoribb a glükóz, galaktóz, mannóz, xilóz; polimerizációs fok 50-300. A hemicellulózok kémiailag stabilabbak, mint a pektin anyagok, nehezebben hidrolizálódnak és kevésbé duzzadnak vízben. A hemicellulózok tartalékanyagként (datolyapálma, datolyaszilva) rakódhatnak le a magsejtek falában. A pektin anyagok és a hemicellulózok kölcsönös átmenetek kapcsolódnak egymáshoz. A sejtfalak mátrixában a poliszacharidokon kívül egy speciális szerkezeti fehérje is jelen van. A cukorarabinóz maradékaihoz kapcsolódik, ezért glikoprotein.
A mátrix poliszacharidok nem csupán kitöltik a cellulóz mikrofibrillumok közötti réseket. Láncaik rendezetten helyezkednek el, és számos kötést képeznek egymással és mikrofibrillákkal, ami jelentősen növeli a sejtfal szilárdságát.
A növények sejtfala gyakran kémiailag módosított. Lignifikáció vagy lignifikáció akkor következik be, amikor a lignin, egy fenolos természetű polimer vegyület, vízben oldhatatlan, lerakódik a mátrixban. A lignifikált sejtfal veszít rugalmasságából, keménysége, nyomószilárdsága meredeken megnő, vízáteresztő képessége csökken. A lignin reagensei a következők: 1) floroglucinol és tömény sósav vagy kénsav (a lignifikált falak cseresznyepiros színt kapnak) és 2) anilin-szulfát, amelynek hatására a lignizált falak citromsárgává válnak. A lignifikáció a xilém (fa) vezető szövetének és a szklerenchyma mechanikai szövetének sejtfalára jellemző.
A suberinizáció vagy suberinizáció a hidrofób polimerek - suberin és viasz - lerakódásának eredményeképpen a sejtfal belső oldalán történik. A Suberin polimer zsírsav-észterek keveréke. A viasz monomerek zsíralkoholok és viasz-észterek. A viasz könnyen eltávolítható szerves oldószerekkel, gyorsan megolvad és kristályokat képez. A suberin egy amorf vegyület, amely nem olvad meg és nem oldódik szerves oldószerekben. A szuberin és a viasz váltakozó párhuzamos rétegeket alkotva belülről film formájában béleli ki a teljes sejtüreget. A szuberin film gyakorlatilag víz- és gázzáró, ezért kialakulása után a sejt általában elpusztul. A dugósodás a parafa belső szövetének sejtfalaira jellemző. A dugós sejtfalon lévő reagens Sudan III, narancsvörös színű.
Az epidermisz integumentális szövetének sejtjeinek külső falai cutinizáción mennek keresztül. A kutin és a viasz váltakozó rétegekben rakódik le a sejtfal külső felületén film - kutikula formájában. A Kutin egy zsírszerű polimer vegyület, kémiai természetében és tulajdonságaiban hasonló a suberinhez. A kutikula megvédi a növényt a víz túlzott elpárolgásától a növény felületéről. Festheti Sudan III reagenssel narancsvörös színben.
A sejtfal mineralizálódása a mátrixban nagy mennyiségű ásványi anyag, leggyakrabban szilícium-dioxid (szilícium-oxid), ritkábban oxalát és kalcium-karbonát lerakódásának köszönhető. Az ásványi anyagok keménységet és törékenységet kölcsönöznek a falnak. A szilícium-dioxid lerakódás a zsurló, a sás és a kalászosok hámsejtjére jellemző. A szilikosodás következtében megszerzett szárak és levelek merevsége védőanyagként szolgál a csigák ellen, emellett jelentősen csökkenti a növények ehetőségét és tápértékét is.
Egyes speciális sejtekben a sejtfal nyálkahártyája figyelhető meg. Ebben az esetben a cellulóz másodlagos fal helyett amorf, erősen hidratált savas poliszacharidok rakódnak le nyálka és gumi formájában, amelyek kémiai természetükben közel állnak a pektin anyagokhoz. A nyálka jól oldódik vízben, nyálkahártya-oldatok képződésével. Ragadós íny, szálakká feszítve. Száradáskor kanos állagúak. A nyálka lerakódásakor a protoplaszt fokozatosan visszaszorul a sejt közepébe, térfogata és a vakuólum térfogata fokozatosan csökken. Végül a sejtüreg teljesen megtelhet nyálkával, és a sejt elhal. Egyes esetekben a nyálka átjuthat az elsődleges sejtfalon a felszínre. A Golgi-készülék a nyálka szintézisében és elválasztásában játszik fő szerepet.
A növényi sejtek által kiválasztott nyálka különféle funkciókat lát el. Tehát a gyökérsapka nyálka kenőanyagként szolgál, hogy megkönnyítse a gyökércsúcs növekedését a talajban. A rovarevő növények nyálkamirigyei (napharmat) felfogó nyálkát választanak ki, amelyhez a rovarok megtapadnak. A maghéj külső sejtjei (len, birsalma, útifű) által kiválasztott nyálka rögzíti a magot a talaj felszínéhez, és megóvja a palántát a kiszáradástól. A nyálkát metilénkék reagenssel kékre festjük.
Az íny felszabadulása általában akkor következik be, amikor a növények megsérülnek. Például a cseresznyében és a szilvában gyakran megfigyelhető az íny szivárgása a törzsek és ágak sérült területeiről. A cseresznyeragasztó egy megszilárdult gumi. Az íny védő funkciót lát el, lefedi a sebet a felületről. A gumik főként a hüvelyesek (akác, tragacanth astragalus) és a rosaceus szilva alcsaládok (cseresznye, szilva, sárgabarack) családjába tartozó fás szárú növényekben képződnek. Az ínyet és a nyálkát a gyógyászatban használják.
A sejtfal a protoplaszt hulladékterméke. A Golgi-készülékben mátrix poliszacharidok, fali glikoprotein, lignin és nyálka képződik. A cellulóz szintézisét, a mikrofibrillumok képződését és orientációját a plazmalemma végzi. A mikrofibrillumok orientációjában fontos szerepet töltenek be a mikrotubulusok, amelyek a plazmalemma közelében, a lerakódott mikrofibrillákkal párhuzamosan helyezkednek el. Ha a mikrotubulusok elpusztulnak, csak izodiametriás sejtek képződnek.
A sejtfal kialakulása a sejtosztódás során kezdődik. Az osztódás síkjában sejtlemez képződik, amely két leánysejtben közös egyetlen réteg. Félig folyékony állagú pektin anyagokból áll; nincs cellulóz. Felnőtt sejtben a sejtlemez megőrződik, de változásokon megy keresztül, ezért mediannak, vagy intercelluláris lemeznek (intercelluláris anyag) nevezik (16. ábra). A középső lemez általában nagyon vékony és szinte megkülönböztethetetlen.
Közvetlenül a sejtlemez kialakulása után a leánysejtek protoplasztjai elkezdik lerakni saját sejtfalukat. Belülről lerakódik mind a sejtlemez felületén, mind más sejtfalak felületén, amelyek korábban az anyasejthez tartoztak. Az osztódást követően a sejt a nyújtó növekedési fázisba lép, ami a sejt intenzív ozmotikus vízfelvételének köszönhető, amely a központi vakuólum kialakulásával és növekedésével jár együtt. A turgornyomás elkezdi feszíteni a falat, de nem törik meg, mivel folyamatosan új mikrofibrillumok és mátrixanyagok rakódnak le benne. Az új anyagrészek lerakódása egyenletesen megy végbe a protoplaszt teljes felületén, így a sejtfal vastagsága nem csökken.
Az osztódó és növekvő sejtek falát elsődlegesnek nevezzük. Sok (60-90%) vizet tartalmaznak. A szárazanyagban a mátrix poliszacharidok (60-70%) dominálnak, a cellulóztartalom nem haladja meg a 30%-ot, és nincs lignin. Az elsődleges fal vastagsága nagyon kicsi (0,1-0,5 mikron).
Sok sejt esetében a sejtfal lerakódása a sejtnövekedés leállásával egyidejűleg leáll. Az ilyen sejteket életük végéig vékony elsődleges fal veszi körül (16. ábra).
Rizs. 16.
Más sejtekben a fal lerakódása akkor is folytatódik, ha a sejt eléri végső méretét. Ebben az esetben a falvastagság nő, és a sejtüreg által elfoglalt térfogat csökken. Ezt a folyamatot másodlagos falvastagodásnak, magát a falat pedig másodlagosnak nevezzük (17. ábra).
A másodlagos fal kiegészítő, főként mechanikai tartófunkciónak tekinthető. Ez a másodlagos fal, amely felelős a fa, a textilszálak, a papír tulajdonságaiért. A másodlagos fal lényegesen kevesebb vizet tartalmaz, mint az elsődleges; dominálnak benne a cellulóz mikrofibrillumok (a szárazanyag tömeg 40-50%-a), amelyek egymással párhuzamosan helyezkednek el. A mátrix poliszacharidjai közül a hemicellulózok (20-30%) jellemzőek, nagyon kevés a pektinanyag. A másodlagos sejtfalak általában lignifikáltak. A nem lignizált másodlagos falakban (len háncsszálak, pamut szőrszálak) a cellulóztartalom elérheti a 95%-ot. A mikrofibrillumok magas tartalma és szigorúan meghatározott orientációja meghatározza a másodlagos falak magas mechanikai tulajdonságait. Gyakran előfordul, hogy a másodlagos lignizált sejtfalú sejtek a másodlagos megvastagodás befejeződése után elpusztulnak.
A lamina median ragasztja a szomszédos sejteket. Ha feloldódik, a sejtfalak elvesztik kapcsolatukat egymással és megszakadnak. Ezt a folyamatot macerációnak nevezik. Elterjedt a természetes maceráció, melynek során a középső tányér pektinanyagai a pektináz enzim segítségével oldható állapotba kerülnek, majd vízzel kimosódnak (körte, dinnye, őszibarack, banán túlérett termései). Gyakran megfigyelhető részleges maceráció, amelyben a medián lemez nem a teljes felületen, hanem csak a sejtek sarkában oldódik fel. A turgornyomás miatt ezeken a helyeken a szomszédos sejtek lekerekednek, ami sejtközi terek kialakulását eredményezi (16. ábra). Az intercelluláris terek egyetlen elágazó hálózatot alkotnak, amelyet vízgőzzel és gázokkal töltenek meg. Így az intercelluláris terek javítják a sejtek gázcseréjét.
A másodlagos fal jellegzetessége a primer falra való egyenetlen lerakódása, aminek következtében nem megvastagodott területek (pórusok) maradnak a szekunder falban. Ha a másodlagos fal nem túl vastag, a pórusok sekély mélyedéseknek tűnnek. Az erős másodlagos falú sejtekben a szakasz pórusai radiális csatornák formájában vannak, amelyek a sejtüregtől az elsődleges falig terjednek. A póruscsatorna alakja szerint kétféle pórus különböztethető meg - egyszerű és szegélyezett (17. ábra).

Rizs. 17. Pórustípusok: A - másodlagos falú sejtek és számos egyszerű pórus; B - egy pár egyszerű pórus; B - egy pár határolt pórus
Egyszerű pórusok esetén a póruscsatorna átmérője teljes hosszában azonos, és keskeny henger alakú. Az egyszerű pórusok a parenchymasejtekre, a háncsra és a farostokra jellemzőek.
Két szomszédos sejt pórusai általában egymással szemben jelennek meg. Ezek a közös pórusok egyetlen csatorna formájúak, vékony szeptum választja el a középső lemeztől és az elsődleges faltól. A szomszédos sejtek szomszédos falainak két pórusából álló halmazt póruspárnak nevezik, és teljes egészében működik. A csatornájukat elválasztó falszakaszt póruszáró filmnek, vagy pórusmembránnak nevezik. Az élő sejtekben a póruszáró filmet számos plazmodezma hatol át (18. ábra).
A plazmodezmák csak a növényi sejtekben rejlenek. Ezek a citoplazma zsinórjai, amelyek áthaladnak a szomszédos sejtek falán. A plazmodezmák száma egy sejtben nagyon nagy - több száztól több tízezerig, általában a plazmodezmákat csoportokban gyűjtik össze. A plazmodezma csatorna átmérője 30-60 nm. Falait plazmalemma béleli, folyamatos a szomszédos sejtek plazmalemmájával. A plazmodesmus közepén egy membránhenger található - a plazmodesmus központi rúdja, amely mindkét sejt endoplazmatikus retikulumának elemeinek membránjaival folyamatos. A központi rúd és a plazmalemma között a csatornában hialoplazma található, amely folyamatos a szomszédos sejtek hialoplazmájával.

Rizs. tizennyolc. Plazmodezmák elektronmikroszkóp alatt (diagram): 1 - hosszanti metszeten; 2 - egy keresztmetszeten; Pl - plasmalemma; TsS - a plazmodezma központi rúdja; ER - az endoplazmatikus retikulum eleme
Így a sejtek protoplasztjai nem különülnek el teljesen egymástól, hanem a plazmodezmák csatornáin keresztül kommunikálnak egymással. Rajtuk keresztül ionok és kismolekulák, valamint hormonális ingerek sejtközi szállítása zajlik. A plazmodezmákon keresztül a növényi szervezet sejtjeinek protoplasztjai egyetlen egészet alkotnak, ezt szimplasztnak nevezik, az anyagoknak a plazmodezmán keresztül történő szállítását pedig szimplasztikusnak nevezik, ellentétben a sejtfalak és sejtközi terek mentén történő apoplasztikus transzporttal.
A bélelt pórusoknál (17. ábra) a sejtfal lerakódása során a csatorna élesen beszűkül, ezért a sejtüregbe nyíló pórus belső nyílása jóval szűkebb, mint a külső, az elsődleges falnak ütköző nyílása. . A szegélyezett pórusok a fa vízvezető elemeinek korán elhaló sejtjeire jellemzőek. Ezekben a póruscsatorna tölcsérszerűen kitágul a zárófólia felé, a másodlagos fal pedig gerinc formájában lóg a csatorna kitágult része fölött, póruskamrát képezve. A szegélyezett pórus elnevezése onnan ered, hogy a belső nyílás felületről nézve kis körnek vagy keskeny résnek tűnik, míg a külső nyílás a belsőt nagyobb átmérőjű kör, ill. szélesebb rés.
A pórusok megkönnyítik a víz és az oldott anyagok sejtről sejtre történő szállítását anélkül, hogy veszélyeztetnék a sejtfal szilárdságát.
Ez a héj szállítási, védelmi és szerkezeti funkciókat lát el. Sok protozoonból hiányzik a sejtfal. A természet az állatokat is megfosztotta ettől az elemtől. A héj a legtöbb prokarióta, archaea és a növényvilág képviselőiben megtalálható.
Bakteriális sejtfal
A héj mureint (peptidoglikánt) tartalmaz. Lehet Gram-pozitív és Gram-negatív is. Az első típusú baktériumok sejtfala kivételesen vastag peptidoglikánréteget tartalmaz. Erősen tapad a membránhoz, és átjárja a lipoteichoin és teichoin savakat. A gram-negatív sejtfal vékonyabb peptidoglikánréteget tartalmaz. A plazmamembrán és közte periplazmatikus tér van. Kívül a héjat egy másik réteg veszi körül. Lipopoliszacharidként kerül bemutatásra. Ez a membrán pirogén endotoxinként működik.
Növényi sejtfal
A cellulóz kulcsfontosságú elemként működik a burkolatban. A sejtfalat a növényvilág legmagasabb képviselőinek legfontosabb jellemzőjének tartják. Ez túlnyomórészt egy összetett polimer mátrix. A fal nélküli sejtet protoplasztnak nevezzük. A héjakban speciális hornyok vannak. A plazmodezmák - citoplazma tubulusok - áthaladnak ezeken a pórusokon. Összekötik a növények egyik sejtfalát a másikkal. Ezek a tubulusok biztosítják az anyagok cseréjét közöttük. Azt kell mondani, hogy a gombák sejtfala sokkal egyszerűbb, mint a növényvilág legmagasabb képviselőinek elemeinek héja.

Kémiai összetétel
A sejt típusától és a szövettől függően változik, amelyben jelen van. Egyes esetekben a kémiai összetétel is megváltozik ugyanazon a héjon belül a protoplaszt körül. A cellulózmolekulák hidrogénkötéseken keresztül kötegeket alkotnak. Ezeket mikrofibrilláknak nevezik. Az összefonódó csomók alkotják a héj vázát. A gombák sejtfala ezen a területen a legtöbb esetben kitint tartalmaz. A mikrofibrillumok a hüvely mátrixában találhatók. Ez viszont különféle vegyi anyagokat tartalmaz. Ezek között a poliszacharidok uralkodnak. Ide tartoznak különösen a pektin anyagok és a hemicellulózok. Tekintsük őket.
Hemicellulóz
Ezek a poliszacharidok egy csoportja. Ezek hexózok és pentózok polimerei - glükóz, galaktóz, mannóz, xilóz stb. A hemicellulózmolekulák, akárcsak a cellulózmolekulák, lánc formájában jelennek meg. Utóbbiaktól azonban rövidebb hosszuk, erős elágazódásuk és kevésbé rendezettségük különbözteti meg őket. Ezeket a láncokat az enzimek könnyebben lebontják és feloldódnak.

Pektin anyagok
Ezeket monoszacharidokból (galaktóz és arabinóz), galakturonsavból (cukor) és metil-alkoholból képzett polimerek képviselik. A pektin molekulák hosszúak. Lehetnek elágazóak vagy lineárisak. Nagyszámú karboxilcsoportot tartalmaznak. Ez lehetővé teszi ezek kombinálását Ca2- és Mg2 + ionokkal. Az eredmény zselatinos, ragadós kalcium- és magnézium-pektát. Ezt követően medián lemezeket alakítanak ki belőlük, amelyekkel az egyik sejtfal a másikhoz kapcsolódik. A fémionok kicserélhetők más kationokra. Ez határozza meg a héjak kationcserélő kapacitását. A pektinek és pektátok sok gyümölcs sejtfalában bővelkednek. Mivel kivonásuk és az azt követő cukor hozzáadása során gélek képződnek, a pektinek zselésítőszerként használják a lekvárgyártás során.

Mátrix
A szénhidrát elemeken kívül tartalmazza a szerkezeti fehérjét, az exteneneint - glikonroteint. Összetételében ez a fehérje közel áll az állatok intercelluláris terében jelenlévő kollagénekhez. A mátrix a héj szárazanyagának körülbelül 60%-át veszi fel. Nemcsak kitölti a mikroszálak közötti réseket, hanem erős kémiai (főleg kovalens és hidrogén) kötéseket képez a cellulózmolekulák és makromolekulák közvetlenül kötegei között. Ez biztosítja a sejtfal szükséges szilárdságát, plaszticitását és rugalmasságát.
Lignin
A héjban a fő kérdőanyagként működik. A lignin aromás alkoholokból álló, el nem ágazó molekulákkal rendelkező polimer. Miután az elemek növekedése leállt, megkezdődik az intenzív lignifikáció. A folyamat során a cellulózmolekulákat impregnálják a polimerrel. A lignin különálló területek - gyűrűk, hálózatok vagy spirálok - formájában halmozódhat fel. Ez különösen a xilém, egy vezető szövet sejtfalára jellemző. A felhalmozódás történhet összefüggő réteg formájában is. A polimer nem csak azokon a területeken rakódik le, ahol a szomszédos sejtekkel érintkezésbe kerül plazmodezma formájában. A lignin a cellulózszálak összetartásával szívós és nagyon merev vázként működik. Növeli a héjak nyomó- és szakítószilárdságát. A lignin további védelmet nyújt a kémiai és fizikai hatásokkal szemben, valamint csökkenti a vízáteresztő képességet. A héj polimertartalma akár 30% is lehet. A lignin inkrusztáció gyakran a falak lignifikációjához vezet. Ez viszont a tartalom halálával jár. A cellulózzal kombinálva a lignin sajátos tulajdonságokat kölcsönöz a fának. Ez viszont sokoldalú építőanyaggá teszi.

Zsíros anyagok
A héjra is lerakhatók. A zsírszerű anyagok közé tartozik a cutin, a viasz és a suberin. Ez utóbbi a sejt belsejéből halmozódik fel. Szinte áthatolhatatlanná teszi az oldatokat és a vizet. Ennek eredményeként a protoplaszt elhal, és a sejt megtelik levegővel. Ezt a folyamatot dugósításnak nevezik. Évelő faültetvények integumentáris szöveteiben figyelhető meg. Az epidermális sejtmembránt viasz és cutin védi. Ezek hidrofób anyagok. Prekurzoraik a citoplazmából választódnak ki a felszínre. Ott polimerizálódnak. A kutinréteget általában poliszacharid elemek (pektin és cellulóz) átitatják. Ez képezi a kutikulát. A viasz gyakran kristályos formában halmozódik fel a növényi elemek felületén (gyümölcsökön, leveleken), és sajátos lepedéket képez. A kutikulával együtt védi a sejtet a fertőzések behatolásától és a különféle károsodásoktól. Ezenkívül csökkentik a víz párolgását.

Mineralizáció
Egyes növények (sás, gabonafélék és mások) epidermális sejtjeinek falában fordul elő. Az ásványi anyagok meglehetősen nagy mennyiségben halmozódnak fel bennük. Elsősorban szilícium-dioxid és kalcium-karbonát található. A mineralizáció során az ültetvények szárai és levelei merevséget, keménységet kapnak és kevésbé károsodnak.
Következtetés
A növények sejtfalának számos funkciója van. Különösen merevséget biztosítanak a mechanikai és szerkezeti alátámasztáshoz, alakzathoz és növekedéshez. A héj megakadályozza a turgor - ozmotikus nyomást. Ez különösen fontos azokban az esetekben, amikor további mennyiségű vizet juttatnak az üzembe.







