Mitokondriális DNS és családtörténet. Humán mitokondriális genetika Mi a mitokondriális DNS
A nukleinsavak szerkezete.
Az N. to.- a sejt univerzális információs makromolekulája, melynek fő funkciója: 1) Örökletes információ tárolása genetikai kód formájában. 2) Örökletes információ reprodukálása ön- vagy DNS-replikációval. 3) Az örökletes információ megvalósítása a fehérje bioszintézis folyamatában. N. to. először kaptak gennysejtek magjából. A kémiai elemzés kimutatta, hogy az n-nek két típusa van: 1) DNS, 2) RNS. A DNS a sejtmagban, a mitokondriumokban és a centriolokban található. Az RNS-ek a sejtmagban, a sejtmagokban, a riboszómákban és a mitokondriumokban találhatók. Kémiai értelemben n to - polimerek, amelyek polinukleotid láncokból állnak, monomer n. to. yav - nukleotid. A DNS-ben 4 típus létezik: A, T, G, C. Az RNS-ben a timint Uracil helyettesíti. Szerkezete szerint az adenin és a guanin purinbázisok
A DNS tulajdonságai és funkciói.
A kémiai elemzés kimutatta, hogy az n-nek két típusa van: 1) DNS, 2) RNS. A DNS a sejtmagban, a mitokondriumokban és a centriolokban található. Az RNS-ek a sejtmagban, a sejtmagokban, a riboszómákban és a mitokondriumokban találhatók. Kémiai értelemben n to - polimerek, amelyek polinukleotid láncokból állnak, monomer n. to. yav - nukleotid. A DNS-ben 4 típus létezik: A, T, G, C. Az RNS-ben a timint Uracil helyettesíti. Szerkezete szerint az adenin és a guanin purinbázisok
1 benzolgyűrű) T, C, U (pirimidin bázisok - 2 benzolgyűrű). A DNS egy hélix, amely két polinukleotid láncból áll. A polinukleotid láncok olyan nukleotidokból állnak, amelyek hidrogénkötésekkel kapcsolódnak egymáshoz, amelyek komplementer párokat alkotnak. A DNS-ben A=T, G=C (Chargaff Rule 1951). A DNS minden szálában lévő nukleotidok úgy kapcsolódnak egymáshoz, hogy az előző nukleotid 5. cukorszénje kapcsolódik a következő nukleotid 3. cukorszénéhez. Ezeknek a kötéseknek köszönhetően a DNS-molekulának két vége van. Az LNC hélix átmérője 2 nm. A hélix egy fordulata 3,4 nm. A nukleotidok közötti távolság 0,34 nm. A hélix minden fordulója 10 bázispárt tartalmaz. A DNS-ben több szerveződési szint létezik: 1) Az elsődleges szerkezet az a sorrend, amelyben a nukleotidok komplementer láncokba rendeződnek. 2) A másodlagos szerkezet a DNS kettős hélixe. 3) harmadlagos - DNS a kromoszómák összetételében.
A mitokondriális DNS szerkezeti jellemzői.
Mitokondriális DNS (mtDNS) - A DNS (a nukleáris DNS-től eltérően) a mitokondriumokban, az eukarióta sejtek organellumáiban lokalizálódik.
A legtöbb vizsgált organizmusban a mitokondriumok csak cirkuláris DNS-molekulákat tartalmaznak, egyes növényekben egyszerre vannak jelen cirkuláris és lineáris molekulák is, és számos protistákban (például csillósokban) csak lineáris molekulák találhatók. Az emlős mitokondriumok jellemzően körkörös DNS-molekulák két-tíz azonos másolatát tartalmazzák. A növényekben minden mitokondrium több, különböző méretű DNS-molekulát tartalmaz, amelyek képesek rekombinációra.A kinetoplasztid rendbe tartozó protistáknál (például tripanoszómák) a mitokondrium egy speciális szakasza (kinetoplaszt) kétféle DNS-molekulát tartalmaz - azonos maxi. gyűrűk (20-50 darab) körülbelül 21 kb hosszúak. és mini-gyűrűk (20 000 - 55 000 darab, kb. 300 fajta, átlagos hossza kb. 1000 bp). Az összes gyűrű egyetlen hálózatba (katenánok) kapcsolódik, amely minden replikációs ciklussal megsemmisül és helyreáll. A maxi-gyűrűk homológok más szervezetek mitokondriális DNS-ével. Minden minigyűrű négy hasonló konzervált régiót és négy egyedi hipervariábilis régiót tartalmaz. A minikörök rövid vezető RNS-molekulákat (guideRNS) kódolnak, amelyek szerkesztik a maxicircle génekből átírt RNS-t. A mitokondriális DNS (mtDNS) a sejtszervecskék - mitokondriumok - genomja. Ezen organellumok endoszimbiotikus eredete meghatározza a mitokondriális genetikai rendszer félig autonóm létezését. Tehát a DNS-szintézis a mitokondriumokban a nukleáris DNS-szintézistől függetlenül megy végbe, és ennek a citoplazmatikus genetikai struktúrának - a mitokondriális kromoszómának - az öröklődése általában szigorúan az anyai vonal mentén történik. Ez okot ad a szerzőknek arra, hogy a mitokondriális gének összességét és minden replikálódó mtDNS-fragmentumot feltételesen különítsenek el a populáció egy külön genetikai erőforrásába - a mitokondriális génállományba. A 60-as években azonosítottak DNS-tartalmú struktúrákat a mitokondriumokban. Az elmúlt negyedszázad során az ember és számos állatfaj mitokondriális genomjának szerkezeti és funkcionális szerveződését részletesen tanulmányozták. A mitokondriális kromoszómát egy körkörös, kétszálú DNS-molekula képviseli, amely a belső mitokondriális membránhoz kapcsolódó, kovalensen zárt szupertekercses formában van jelen az organellumban. Minden organellum 1-8 DNS-molekulát tartalmaz, ami sejtenként 1000-8000 kópia. Általános szabály, hogy egy szervezetnek egyetlen formája van az mtDNS-nek, pl. az egyik haplotípus az anyai vonalból öröklődött.
RNS típusok a sejtekben.
A sejtekben háromféle RNS található: 1) I-RNS (hírvivő vagy hírvivő RNS).
2) R-RNS (riboszomális RNS).
3) T-RNS (transzfer RNS)
Messenger RNS - szintetizálódik és átíródik DNS-be, és információt hordoz a fehérjeszintézishez. Az R-RNS és a T-RNS szintetizálódik a sejtmag magjában. A nucleolus a kromoszómák olyan régiója, amelyben műholdak találhatók. A nukleoláris DNS tartalmazza azokat a géneket, amelyeken az R-RNS és a T-RNS szintetizálódik. Az rRNS-ek a riboszómákban találhatók (a kis és nagy alegységben). Célja: az AK kis alegységén keresztül ATP-n keresztül kapcsolódik a T-RNS-hez. A DNS és az RNS közötti különbség: 1) Az RNS egyetlen láncból áll. 2) Az RNS-ben a cukor ribóz. 3) Az RNS rövidebb, mint a DNS. 4) A T-RNS harmadlagos szerkezetű. Mátrix (információs) RNS - RNS, amely közvetítőként szolgál a DNS-ben kódolt információk riboszómákba, az élő szervezet fehérjeit szintetizáló molekuláris gépeibe történő átvitelében. Az mRNS kódoló szekvencia határozza meg a fehérje polipeptidláncának aminosavszekvenciáját.
Transport (tRNS) - kicsi, körülbelül 80 nukleotidból áll, konzervatív tercier szerkezetű molekulákból. Specifikus aminosavakat szállítanak a peptidkötés szintézisének helyére a riboszómában. Mindegyik tRNS tartalmaz egy aminosav kapcsolódási helyet és egy antikodont az mRNS kodonok felismeréséhez és kapcsolódásához. A riboszómális RNS (rRNS) a riboszómák katalitikus komponense. Az eukarióta riboszómák négyféle rRNS-molekulát tartalmaznak: 18S, 5.8S, 28S és 5S. Az rRNS négy típusa közül három a sejtmagban szintetizálódik. A citoplazmában a riboszómális RNS-ek a riboszómális fehérjékkel egyesülve egy riboszómának nevezett nukleoproteint képeznek. A riboszóma az mRNS-hez kapcsolódik, és szintetizálja a fehérjét. Az rRNS az eukarióta sejt citoplazmájában található RNS 80%-át teszi ki
Fő cikk: Mitokondriális DNS
A mátrixban található mitokondriális DNS egy zárt, körkörös, kétszálú molekula, emberi sejtekben 16569 nukleotidpár méretű, ami körülbelül 105-ször kisebb, mint a sejtmagban lokalizált DNS. A mitokondriális DNS általában 2 rRNS-t, 22 tRNS-t és 13 légzőlánc-enzim alegységet kódol, ami a benne található fehérjék legfeljebb felét teszi ki. Különösen a mitokondriális genom szabályozása alatt hét ATP szintetáz alegység, három citokróm oxidáz alegység és egy ubikinol citokróm alegység kódolódik. Val vel-reduktázok. Ebben az esetben a nehezebb (külső) DNS-láncból minden fehérje, egy kivételével két riboszómális és hat transzport RNS, a könnyebb (belső) láncból pedig 14 másik tRNS és egy fehérje íródik át.
Ennek fényében a növényi mitokondriális genom sokkal nagyobb, és elérheti a 370 000 nukleotidpárt is, ami körülbelül 20-szor nagyobb, mint a fent leírt humán mitokondriális genom. A gének száma itt is körülbelül 7-szer nagyobb, ami azzal jár, hogy a növényi mitokondriumokban további elektrontranszport utak jelennek meg, amelyek nem kapcsolódnak az ATP szintézishez.
A mitokondriális DNS interfázisban replikálódik, ami részben szinkronban van a sejtmagban lévő DNS-replikációval. A sejtciklus során a mitokondriumok szűküléssel kettéosztódnak, amelyek kialakulása a belső mitokondriális membránon található gyűrűs barázdával kezdődik. A mitokondriális genom nukleotidszekvenciájának részletes vizsgálata lehetővé tette annak megállapítását, hogy az univerzális genetikai kódtól való eltérések nem ritkák az állatok és gombák mitokondriumaiban. Így a humán mitokondriumokban a standard kódban az izoleucin helyett a TAT kodon a metionin aminosavat kódolja, az általában arginint kódoló TCT és TCC kodonok stopkodonok, és az ACT kodon, amely a standardban stopkodon. kód, a metionin aminosavat kódolja. Ami a növényi mitokondriumokat illeti, úgy tűnik, hogy univerzális genetikai kódot használnak. A mitokondriumok másik jellemzője a tRNS kodonfelismerés jellemzője, amely abból áll, hogy egy ilyen molekula nem egy, hanem három vagy négy kodont képes felismerni egyszerre. Ez a tulajdonság csökkenti a harmadik nukleotid jelentőségét a kodonban, és ahhoz a tényhez vezet, hogy a mitokondriumok kisebb tRNS-típusokat igényelnek. Ebben az esetben csak 22 különböző tRNS elegendő.
A saját genetikai apparátussal rendelkező mitokondriumnak saját fehérjeszintetizáló rendszere is van, melynek jellemzője az állatok és gombák sejtjeiben nagyon kicsi riboszómák, amelyeket 55S-es ülepedési együttható jellemez, ami még a 70S-riboszómákénál is alacsonyabb. prokarióta típusú. Ugyanakkor két nagy riboszomális RNS is kisebb, mint a prokariótákban, és a kis rRNS teljesen hiányzik. Ezzel szemben a növényi mitokondriumokban a riboszómák méretükben és szerkezetükben jobban hasonlítanak a prokariótákhoz.
Mitokondriális fehérjék[szerkesztés | forrás szerkesztése]
A mitokondriális mRNS-ből lefordított fehérjék száma, amelyek nagy enzimkomplexek alegységeit alkotják, korlátozott. A fehérjék jelentős része a sejtmagban kódolódik, és a citoplazmatikus 80S riboszómákon szintetizálódik. Különösen egyes fehérjék képződnek ilyen módon - elektronhordozók, mitokondriális transzlokázok, a fehérjék mitokondriumokba történő szállításának összetevői, valamint a mitokondriális DNS transzkripciójához, transzlációjához és replikációjához szükséges tényezők. Ugyanakkor az ilyen fehérjék N-terminálisán speciális szignálpeptidek találhatók, amelyek mérete 12 és 80 aminosav között változik. Ezek a helyek amfifil fürtöket képeznek, specifikus érintkezést biztosítanak a fehérjéknek a külső membránon lokalizált mitokondriális felismerő receptorok kötődoménjeivel. A külső mitokondriális membránba ezek a fehérjék részben kibontott állapotban a chaperon fehérjékkel (különösen a hsp70-nel) együtt szállítódnak. A külső és belső membránon keresztül, érintkezési pontjain átjutva a mitokondriumokba kerülő fehérjék ismét a chaperonokhoz kötődnek, de saját mitokondriális eredetűek, amelyek felveszik a membránokon áthaladó fehérjét, elősegítik annak visszahúzódását a mitokondriumba, és szabályozzák is. a polipeptidlánc megfelelő feltekeredésének folyamata. A chaperonok többsége ATPáz aktivitással rendelkezik, aminek eredményeként mind a fehérjék mitokondriumba szállítása, mind funkcionálisan aktív formáinak kialakulása energiafüggő folyamat.
Bevezetés
Negyedszázad telt el a DNS-molekulák felfedezése óta a mitokondriumokban, mire nem csak a molekuláris biológusok és citológusok, hanem a genetika, az evolúciós kutatók, valamint a paleontológusok és a törvényszéki tudósok iránt is érdeklődni kezdtek. Ekkora érdeklődést A. Wilson, a Kaliforniai Egyetem munkatársa munkája váltott ki. 1987-ben publikálta a mitokondriális DNS összehasonlító elemzésének eredményeit, amelyet öt kontinensen élő összes emberi faj különböző etnikai csoportjainak 147 képviselőjétől vettek. Az egyedi mutációk típusa, elhelyezkedése és száma szerint megállapították, hogy az összes mitokondriális DNS egy ősi nukleotidszekvenciából származott divergenciával. Az áltudományos sajtóban ezt a következtetést rendkívül leegyszerűsítve értelmezték - az egész emberiség egyetlen nőtől származott, akit mitokondriális Évának neveztek (mivel a lányok és a fiúk is csak az anyjuktól kapják a mitokondriumokat), aki körülbelül 200 ezer évvel ezelőtt élt Északkelet-Afrikában. Újabb 10 év elteltével sikerült megfejteni a neandervölgyi maradványaiból izolált mitokondriális DNS-részletet, és 500 ezer évvel ezelőttre becsülni az ember és a neandervölgyi utolsó közös őse létezésének idejét.
Napjainkban a humán mitokondriális genetika intenzíven fejlődik mind a populációban, mind az orvosi szempontból. Összefüggést állapítottak meg számos súlyos örökletes betegség és a mitokondriális DNS hibája között. Az öregedéssel összefüggő genetikai változások a legkifejezettebbek a mitokondriumokban. Mi az a mitokondriális genom, amely méretét, alakját és genetikai kapacitását tekintve különbözik az emberben és más állatokban a növények, gombák és protozoonok genomjától? Mi a szerepe, hogyan működik, és hogyan keletkezett a mitokondriális genom általában különböző taxonokban, és konkrétan az emberben? Erről a „kicsi és legszerényebb” esszémben lesz szó.
A mitokondriális mátrix a DNS mellett saját riboszómákat is tartalmaz, amelyek sok jellemzőben különböznek az endoplazmatikus retikulum membránjain található eukarióta riboszómáktól. A mitokondriális riboszómák azonban az összetételüket alkotó összes fehérje legfeljebb 5%-át alkotják. A mitokondriumok szerkezeti és funkcionális komponenseit alkotó fehérjék többségét a nukleáris genom kódolja, az endoplazmatikus retikulum riboszómáin szintetizálódnak, és csatornáin keresztül jutnak el a gyülekezési helyre. Így a mitokondriumok két genom és két transzkripciós és transzlációs berendezés együttes erőfeszítésének eredményei. A mitokondriális légzőlánc egyes alegység-enzimei különböző polipeptidekből állnak, amelyek egy részét a nukleáris, másokat a mitokondriális genom kódol. Például az élesztőben az oxidatív foszforiláció kulcsenzime, a citokróm-c-oxidáz három mitokondriumban kódolt és szintetizált alegységből, valamint a sejtmagban kódolt és a citoplazmában szintetizált négy alegységből áll. A legtöbb mitokondriális gén expresszióját bizonyos nukleáris gének szabályozzák.
A mitokondriumok eredetének szimbiotikus elmélete
A mitokondriumok és a növényi plasztidok intracelluláris endoszimbionta baktériumokból való eredetére vonatkozó hipotézist R. Altman vetette fel még 1890-ben. A biokémia, citológia, genetika és molekuláris biológia fél évszázaddal ezelőtti gyors fejlődésének évszázada során a hipotézis tényszerűbb anyagokon alapuló elméletté nőtte ki magát . Lényege a következő: a fotoszintetikus baktériumok megjelenésével a Föld légkörében felhalmozódott oxigén - anyagcseréjük mellékterméke. Koncentrációjának növekedésével az anaerob heterotrófok élete bonyolultabbá vált, és egy részük az anoxikus fermentációról oxidatív foszforilációra váltott, hogy energiát nyerjen. Az ilyen aerob heterotrófok az anaerob baktériumoknál nagyobb hatékonysággal képesek lebontani a fotoszintézis eredményeként keletkező szerves anyagokat. A szabadon élő aerobok egy részét az anaerobok befogták, de nem „emésztették meg”, hanem energiaállomásokként, mitokondriumokként tárolták. A mitokondriumokat nem szabad rabszolgának tekinteni, hogy ATP-molekulákkal látják el azokat a sejteket, amelyek nem képesek légzésre. Ők inkább „lények”, akik még a proterozoikumban is megtalálták a legjobb menedéket maguknak és utódaiknak, ahol a legkevesebb erőfeszítést tudják ráfordítani anélkül, hogy megeszik őket.
Számos tény szól a szimbiotikus elmélet mellett:
A mitokondriumok és a szabadon élő aerob baktériumok mérete és alakja egybeesik; mindkettő cirkuláris DNS-molekulákat tartalmaz, amelyek nem kapcsolódnak hisztonokhoz (ellentétben a lineáris nukleáris DNS-sel);
A mitokondriumok riboszómális és transzport RNS-ei nukleotidszekvenciában különböznek a nukleáris szekvenciáktól, miközben meglepő hasonlóságot mutatnak néhány aerob gram-negatív eubaktérium analóg molekuláival;
A mitokondriális RNS polimerázokat, bár a sejtmagban kódolják, a rifampicin gátolja, akárcsak a bakteriálisakat, és az eukarióta RNS polimerázok érzéketlenek erre az antibiotikumra;
A mitokondriumokban és a baktériumokban a fehérjeszintézist ugyanazok az antibiotikumok gátolják, amelyek nem befolyásolják az eukarióta riboszómákat;
A belső mitokondriális membrán és a bakteriális plazmalemma lipidösszetétele hasonló, de nagyon különbözik a külső mitokondriális membránétól, amely homológ az eukarióta sejtek más membránjaival;
A belső mitokondriális membrán által alkotott kriszták számos prokarióta mezoszomális membránjának evolúciós analógjai;
Mostanáig túléltek olyan organizmusok, amelyek a köztes formákat utánozzák a baktériumokból (primitív amőba) történő mitokondriumok kialakulásához vezető úton Pelomyxa nem tartalmaz mitokondriumokat, de mindig tartalmaz endoszimbiotikus baktériumokat).
Van egy olyan elképzelés, hogy az eukarióták különböző birodalmainak eltérő ősei voltak, és a baktériumok endoszimbiózisa az élő szervezetek fejlődésének különböző szakaszaiban alakult ki. Ezt bizonyítja a protozoonok, gombák, növények és magasabb rendű állatok mitokondriális genomjának szerkezeti különbségei is. De minden esetben a promitokondriumokból származó gének nagy része a sejtmagba került, esetleg mobil genetikai elemek segítségével. Ha az egyik szimbionta genomjának egy része bekerül a másik genomjába, a szimbionták integrációja visszafordíthatatlanná válik. Az új genom olyan metabolikus utakat hozhat létre, amelyek olyan hasznos termékekhez vezetnek, amelyeket egyik partner sem képes önmagában szintetizálni. Így a szteroid hormonok szintézise a mellékvesekéreg sejtjeiben egy összetett reakciólánc, amelyek egy része a mitokondriumokban, más része pedig az endoplazmatikus retikulumban fordul elő. A promitokondriumok génjeinek rögzítése után a mag képes volt megbízhatóan ellenőrizni a szimbionta funkcióit. A sejtmag kódolja a mitokondriumok külső membránjának összes fehérjét és lipidszintézisét, a mátrix legtöbb fehérjét és az organellumok belső membránját. A legfontosabb, hogy a sejtmag az mtDNS replikáció, transzkripció és transzláció enzimeit kódolja, ezáltal szabályozza a mitokondriumok növekedését és szaporodását. A szimbiózisban lévő partnerek növekedési ütemének megközelítőleg azonosnak kell lennie. Ha a gazda gyorsabban növekszik, akkor minden generációval csökken az egyedre jutó szimbionták száma, és a végén megjelennek olyan leszármazottak, amelyeknek nincs mitokondriumuk. Tudjuk, hogy egy szexuálisan szaporodó szervezet minden sejtje sok mitokondriumot tartalmaz, amelyek a DNS-üket replikálják a gazdaosztályok között. Ez biztosítja, hogy minden leánysejt megkapja a mitokondriális genom legalább egy másolatát.
A sejtmag szerepe a mitokondriális biogenezisben
Egy bizonyos típusú mutáns élesztőben nagy deléció található a mitokondriális DNS-ben, ami a fehérjeszintézis teljes leállásához vezet a mitokondriumokban; ennek következtében ezek az organellumok nem képesek ellátni funkciójukat. Mivel az ilyen mutánsok kis telepeket képeznek, ha alacsony glükóztartalmú táptalajon termesztik őket, ún citoplazmatikus mutantamiaprócska.
Bár a kisméretű mutánsok nem termelnek mitokondriális fehérjeszintézist, és ezért nem képeznek normális mitokondriumokat, ezek a mutánsok mégis tartalmaznak promitokondriumok, amelyek bizonyos mértékig hasonlítanak a közönséges mitokondriumokhoz, normál külső membránnal és egy belső membránnal gyengén fejlett cristae-kkal rendelkeznek. A promitokondriumok számos sejtmaggének által kódolt és citoplazmatikus riboszómákon szintetizált enzimet tartalmaznak, beleértve a DNS- és RNS-polimerázokat, a citromsavciklus összes enzimét, és számos fehérjét, amelyek a belső membránt alkotják. Ez egyértelműen mutatja a nukleáris genom domináns szerepét a mitokondriális biogenezisben.
Érdekes megjegyezni, hogy bár az elveszett DNS-fragmensek a mitokondriális genom 20-tól több mint 99,9%-át teszik ki, a kisméretű mutánsokban a mitokondriális DNS teljes mennyisége mindig ugyanazon a szinten marad, mint a vad típusban. Ez annak köszönhető, hogy a DNS-amplifikáció még kevéssé tanulmányozott folyamata, amelynek eredményeként olyan DNS-molekula képződik, amely azonos helyű tandem ismétlődésekből áll, és mérete megegyezik egy normál molekulával. Például egy kis mutáns mitokondriális DNS-e, amely megtartja a vad típusú DNS-nukleotidszekvencia 50%-át, két ismétlődésből áll, míg egy molekula, amely csak 0,1% vad típusú genomot állítanak elő a maradék fragmentum 1000 kópiájából. Így a kisméretű mutánsok segítségével nagy mennyiségű mitokondriális DNS-régiót lehet nyerni, amelyekről azt mondhatjuk, hogy maga a természet klónozta.
Bár az organellumok biogenezisét főként maggének szabályozzák, maguk az organellumok is – egyes adatokból ítélve – valamilyen szabályozó visszacsatoló hatást fejtenek ki; mindenesetre ez a helyzet a mitokondriumokkal. Ha az ép sejtek mitokondriumában a fehérjeszintézis blokkolva van, akkor a DNS, RNS és fehérjék mitokondriális szintézisében részt vevő enzimek feleslegben kezdenek képződni a citoplazmában, mintha a sejt megpróbálná leküzdeni a blokkolószer hatását. De bár a mitokondriumból származó jelek létezése kétségtelen, természete még mindig nem ismert.
Számos okból kifolyólag a mitokondriális biogenezis mechanizmusait a legtöbb esetben kultúrákban tanulmányozzák. Saccharomyces carlsbergensis(sörélesztő és S. cerevisiae(sütőélesztő). Először is, amikor glükózon termesztik, ezek az élesztőgombák egyedülálló képességet mutatnak arra, hogy csak a glikolízisnek köszönhetően létezzenek, vagyis a mitokondriumok funkciója nélkül. Ez lehetővé teszi a mitokondriális és nukleáris DNS azon mutációinak tanulmányozását, amelyek megakadályozzák ezen organellumok fejlődését. Az ilyen mutációk szinte minden más organizmusban halálosak. Másodszor, az élesztőgomba - egyszerű egysejtű eukarióták - könnyen tenyészthető, és biokémiai kutatásoknak vetik alá. Végül az élesztő haploid és diploid fázisban is képes szaporodni, általában ivartalanul, bimbózás útján (aszimmetrikus mitózis). De az élesztőben szexuális folyamat is zajlik: időről időre két haploid sejt összeolvad, diploid zigótát képezve, amely vagy mitózissal osztódik, vagy meiózison megy keresztül, és ismét haploid sejteket hoz létre. A kísérlet során az ivartalan és ivaros szaporodás váltakozásának szabályozásával sokat megtudhatunk a mitokondriumok működéséért felelős génekről. Ezekkel a módszerekkel különösen azt lehet kideríteni, hogy az ilyen gének a nukleáris DNS-ben vagy a mitokondriális DNS-ben lokalizálódnak-e, mivel a mitokondriális gének mutációi nem öröklődnek a nukleáris gének öröklődését szabályozó Mendel-törvények szerint.
Mitokondriális transzportrendszerek
A mitokondriumokban és a kloroplasztiszokban található fehérjék nagy része a citoszolból importálódik ezekbe az organellákba. Ez két kérdést vet fel: hogyan irányítja a sejt a fehérjéket a megfelelő organellumokhoz, és hogyan jutnak be ezek a fehérjék?
Részleges választ kaptunk az enzim kis alegységének (S) a kloroplaszt sztrómájába történő transzportjának vizsgálatával. ribulóz-1,5-biszfoszfát-karboxiaknák. Ha egy egysejtű alga citoplazmájából izolált mRNS Chlamydomonas vagy borsólevélből, templátként egy in vitro fehérjeszintézis rendszerbe juttatva, akkor a sok eredményül kapott fehérje közül az egyik specifikus anti-S antitesthez kötődik. Az in vitro szintetizált S-fehérjét ppo-S-nek nevezik, mert körülbelül 50 aminosavval nagyobb, mint a normál S-fehérje. A pro-S fehérje ép kloroplasztiszokkal való inkubációja során behatol az organellumokba, és ott peptidáz hatására S-proteinné alakul. Az S fehérje ezután a kloroplasztisz riboszómákon szintetizált ribulóz-1,5-biszfoszfát-karboxiláz nagy alegységéhez kötődik, és aktív enzimet képez vele a kloroplasztisz stromában.
Az S-fehérje transzfer mechanizmusa nem ismert. Úgy gondolják, hogy a pro-S a kloroplaszt külső membránján vagy a külső és belső membrán találkozásánál elhelyezkedő receptorfehérjéhez kötődik, majd energiaigényes folyamat során transzmembrán csatornákon keresztül a stromába kerül.
Hasonló módon a fehérjék a mitokondriumokba szállítódnak. Ha a tisztított élesztő mitokondriumokat újonnan szintetizált radioaktív élesztőfehérjéket tartalmazó sejtkivonattal inkubálják, megfigyelhető, hogy a nukleáris genom által kódolt mitokondriális fehérjék elkülönülnek a nem mitokondriális citoplazmatikus fehérjéktől, és szelektíven beépülnek a mitokondriumokba, akárcsak egy ép sejtben. . Ugyanakkor a külső és belső membrán, a mátrix és a membránközi tér fehérjéi utat találnak a megfelelő mitokondriális kompartmentbe.
A belső membránba, mátrixba és membránközi térbe szánt újonnan szintetizált fehérjék közül sok N-terminálisán van egy vezető peptid, amelyet a mátrixban található specifikus proteáz a szállítás során lehasít. A fehérjék ebbe a három mitokondriális részébe történő átviteléhez a belső membránon keresztül létrejövő elektrokémiai proton gradiens energiája szükséges. A külső membrán fehérjetranszportjának mechanizmusa eltérő: ebben az esetben nincs szükség sem energiafelhasználásra, sem a hosszabb prekurzor fehérje proteolitikus hasítására. Ezek és más megfigyelések azt sugallják, hogy a mitokondriális fehérjék mind a négy csoportja a következő mechanizmussal kerül az organellumba: feltételezzük, hogy a külső membránra szánt fehérjék kivételével minden fehérje bekerül a belső membránba egy energia hatására. -igényes folyamat, amely a külső és a belső membrán érintkezési pontjain megy végbe. Nyilvánvalóan a fehérje membránba való kezdeti beépülése után proteolitikus hasításon megy keresztül, ami konformációjának megváltozásához vezet; attól függően, hogy a konformáció hogyan változik, a fehérje vagy rögzül a membránban, vagy "kinyomódik" a mátrixba vagy a membránközi térbe.
A fehérjék átvitele a mitokondriumok és a kloroplasztiszok membránjain keresztül elvileg hasonló az endoplazmatikus retikulum membránjain keresztül történő átvitelükhöz. Itt azonban van néhány lényeges különbség. Először is, amikor a fehérje a mátrixba vagy a stromába kerül, az organellum külső és belső membránján is áthalad, míg az endoplazmatikus retikulum lumenébe kerülve a molekulák csak egy membránon haladnak át. Ezenkívül a fehérjék átvitele a retikulumba a mechanizmus segítségével történik irányított kiválasztás(vektoriális kisülés) - akkor kezdődik, amikor a fehérje még nem szállt le teljesen a riboszómáról (kotranszlációs import),és a mitokondriumokba és a kloroplasztiszokba történő átvitel a fehérjemolekula szintézisének teljes befejeződése után következik be (transzláció utáni import).
E különbségek ellenére mindkét esetben a sejt prekurzor fehérjéket szintetizál, amelyek egy szignálszekvenciát tartalmaznak, amely meghatározza, hogy ez a fehérje melyik membránba kerül. Nyilvánvalóan sok esetben ez a szekvencia a transzportfolyamat befejezése után leszakad a prekurzor molekuláról. Egyes fehérjék azonban azonnal szintetizálódnak végső formájukban. Úgy gondolják, hogy ilyen esetekben a szignálszekvencia a kész fehérje polipeptidláncában található. A jelszekvenciák még mindig rosszul ismertek, de valószínűleg többféle ilyen szekvenciának kell lennie, amelyek mindegyike meghatározza egy fehérjemolekula átvitelét a sejt egy meghatározott területére. Például egy növényi sejtben azon fehérjék egy része, amelyek szintézise a citoszolban kezdődik, azután a mitokondriumokba, mások a kloroplasztiszokhoz, mások a peroxiszómákba, mások az endoplazmatikus retikulumba szállítódnak. A fehérjék helyes intracelluláris eloszlásához vezető összetett folyamatok csak most válnak világossá.
A nukleinsavak és fehérjék mellett lipidekre is szükség van az új mitokondriumok felépítéséhez. A kloroplasztiszokkal ellentétben a mitokondriumok lipidjeik nagy részét kívülről kapják. Állati sejtekben az endoplazmatikus retikulumban szintetizált foszfolipidek specifikus fehérjék révén a külső mitokondriális membránba kerülnek, majd beépülnek a belső membránba; ez feltehetően a két membrán érintkezési pontján történik. A lipid bioszintézis fő reakciója, amelyet maguk a mitokondriumok katalizálnak, a foszfatidsav átalakulása foszfolipid cardiolipinné, amely főként a belső mitokondriális membránban található, és az összes lipid körülbelül 20%-át teszi ki.
A mitokondriális genomok mérete és alakja
Eddig több mint 100 különböző mitokondriális genomot olvastak le. A mitokondriális DNS-ben lévő génjeik készlete és száma, amelyek nukleotidszekvenciája teljesen meghatározott, nagymértékben változik a különböző állat-, növény-, gomba- és protozoonfajokban. A legtöbb gént a flagellated protozoon mitokondriális genomjában találták meg Rectinomo-nas americana- 97 gén, beleértve az összes fehérjét kódoló gént, amely más szervezetek mtDNS-ében található. A legtöbb magasabb rendű állatban a mitokondriális genom 37 gént tartalmaz: 13 gént a légzőlánc fehérjéihez, 22 tRNS-éhez és kettő rRNS-éhez (a nagy riboszóma alegységhez, a 16S rRNS-hez és a kis 12S rRNS-hez). A növényekben és a protozoonokban, ellentétben az állatokkal és a legtöbb gombával, a mitokondriális genom néhány olyan fehérjét is kódol, amelyek ezeknek az organellumoknak a riboszómáit alkotják. A templát polinukleotid szintézis kulcsenzimei, mint például a DNS-polimeráz (replikálódik a mitokondriális DNS-t) és az RNS-polimeráz (a mitokondriális genomot átírja), a sejtmagban kódolódnak, és a citoplazmatikus riboszómákon szintetizálódnak. Ez a tény jelzi a mitokondriumok viszonylagos autonómiáját az eukarióta sejt összetett hierarchiájában.
A különböző fajok mitokondriális genomjai nemcsak a gének készletében, elhelyezkedésük és kifejeződésük sorrendjében, hanem a DNS méretében és alakjában is különböznek egymástól. A ma leírt mitokondriális genomok túlnyomó többsége cirkuláris szupertekercselés, kétszálú DNS-molekula. Egyes növényekben a gyűrűs formák mellett lineárisak is vannak, egyes protozoonokban, például csillósokban, csak lineáris DNS-t találtak a mitokondriumokban.
Általában minden mitokondrium genomjának több másolatát tartalmazza. Tehát az emberi májsejtekben körülbelül 2 ezer mitokondrium található, és mindegyikben 10 azonos genom található. Az egér fibroblasztokban 500 mitokondrium van, amelyek egyenként két genomot tartalmaznak, és az élesztősejtekben S.cerevisiae- legfeljebb 22 mitokondrium négy genommal.
DIV_ADBLOCK764">
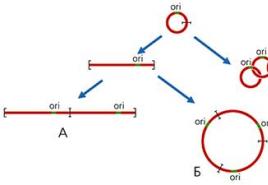
 2. ábra. Lineáris (A), cirkuláris (B), láncos (C) mtDNS oligomerek képződésének sémája. ori - a DNS-replikáció origójának régiója.
2. ábra. Lineáris (A), cirkuláris (B), láncos (C) mtDNS oligomerek képződésének sémája. ori - a DNS-replikáció origójának régiója.
A különböző organizmusok mitokondriális genomjának mérete a maláriás plazmódiumban található kevesebb mint 6 ezer bázispártól (két rRNS gén mellett mindössze három fehérjét kódoló gént tartalmaz) a szárazföldi növényekben található több százezer bázispárig terjed (pl. , ban ben Arabidopsis thaliana a keresztesvirágúak családjából 366924 bázispár). Ugyanakkor a magasabb rendű növények mtDNS méretében még egy családon belül is 7-8-szoros eltéréseket találunk. A gerincesek mtDNS-ének hossza kissé eltér: emberben - 16569 bázispár, sertésben - 16350, delfinekben - 16330, karmos békákban Xenopus laevis- 17533, ponty - 16400. Ezek a genomok a gén lokalizációját tekintve is hasonlóak, legtöbbjük végek között helyezkedik el; egyes esetekben még át is fedik egymást, általában egy nukleotiddal, így az egyik gén utolsó nukleotidja az első a következőben. A gerincesekkel ellentétben a növényekben, gombákban és protozoonokban az mtDNS a nem kódoló szekvenciák 80%-át tartalmazza. A különböző fajoknál a gének sorrendje a mitokondriumok genomjában eltérő.
A reaktív oxigénfajták magas koncentrációja a mitokondriumokban és a gyenge javítórendszer nagyságrenddel növeli az mtDNS mutációk gyakoriságát a nukleárishoz képest. Az oxigéngyökök a C®T (citozin dezaminálása) és a G®T (guanin oxidatív károsodása) specifikus szubsztitúcióit okozzák, aminek következtében az mtDNS valószínűleg gazdag AT-párokban. Ezenkívül minden mtDNS-nek van egy érdekes tulajdonsága - a nukleáris és prokarióta DNS-sel ellentétben nem metiláltak. Ismeretes, hogy a metiláció (a nukleotidszekvencia ideiglenes kémiai módosítása a DNS kódoló funkciójának megsértése nélkül) a programozott géninaktiválás egyik mechanizmusa.
A DNS-molekulák mérete és szerkezete az organellumokban
Szerkezet | Súly, millió dalton | Megjegyzések |
||
|
ohon drya | Állatok | Gyűrű | Minden egyes fajban minden molekula azonos méretű |
|
magasabb ra nyögve | Gyűrű | Váltakozik | Minden vizsgált fajnak különböző méretű, kör alakú DNS-ei vannak, amelyekben a genetikai információ teljes tartalma fajtól függően 300-1000 millió dalton tömegnek felel meg. |
|
Gomba: Protozoa | Gyűrű Gyűrű Gyűrű Lineáris | |||
|
Klór fizetés tűzhely | Hínár | Gyűrű Gyűrű | ||
magasabb növények | Gyűrű | Minden egyes fajban csak egy molekulát találtak. |
Az organellum DNS relatív mennyisége egyes sejtekben és szövetekben
szervezet | szövet ill sejttípus | Mol-l DNS/organellum- | a szervek száma- nell be ketrec | Az organellum DNS aránya az összesben sejt DNS, % |
|
|
ohon drya | |||||
L vonalsejtek | |||||
Tojás | |||||
|
Klór fizetés tűzhely | Vegetatív diploid sejtek | ||||
Kukorica |
A mitokondriális genom működése
Mi a különleges az emlős mitokondriumokban a DNS-replikáció és transzkripció mechanizmusában?
Az mtDNS komplementer" href="/text/category/komplementarij/" rel="bookmark">komplementer láncai fajlagos sűrűségükben jelentősen eltérnek egymástól, mivel egyenlőtlen mennyiségű „nehéz” purin és „könnyű” pirimidin nukleotidot tartalmaznak. - H (nehéz - nehéz) és L (könnyű - könnyű) láncnak nevezik. Az mtDNS molekula replikációjának kezdetén kialakul az úgynevezett D-hurok (az angolból. Displacement loop - displacement loop). Ez az elektronikus mikroszkópban látható szerkezet kétszálú és egyszálú (a H-lánc egy része eltávolodott) szakaszokból áll. A kétszálú szakaszt az L-lánc egy része és egy újonnan szintetizált DNS-fragmens, amely komplementer vele 450-650 nukleotid hosszúsággal (a szervezet típusától függően), a ribonukleotid primer 5"-végével, amely megfelel a H-lánc szintézis kezdőpontjának (oriH). Az L-lánc szintézise csak akkor kezdődik meg, amikor a leány-H-lánc eléri az ori L pontot. Ennek oka az a tény, hogy az L-lánc replikációjának iniciációs tartománya a DNS-szintézis enzimek számára csak egyetlen egy- sodrott állapotban, és ezért csak egy csavaratlan kettős hélixben a H-lánc szintézisében. Így az mtDNS leányszálai folyamatosan és aszinkron módon szintetizálódnak (3. ábra).
3. ábra. Az mtDNS replikáció sémája emlősökben. Először a D-hurok jön létre, majd a leány-H-lánc szintetizálódik, majd megkezdődik a leány-L-lánc szintézise.
A 16S rRNS gén vége (4. ábra). 10-szer több ilyen rövid átirat van, mint hosszú. Az érés (feldolgozás) eredményeként belőlük 12S rRNS és 16S rRNS képződik, amelyek részt vesznek a mitokondriális riboszómák, valamint a fenilalanin és a valin tRNS képzésében. A fennmaradó tRNS-eket kivágják a hosszú transzkriptumokból, és transzlált mRNS-ek keletkeznek, amelyek 3'-végeihez poliadenil-szekvenciák kapcsolódnak, amelyek 5'-végei nincsenek lezárva, ami szokatlan az eukariótáknál. Illesztés (fúzió) nem fordul elő, mivel az emlős mitokondriális gének egyike sem tartalmaz intronokat.
4. ábra. 37 gént tartalmazó humán mtDNS transzkripciója. Az összes transzkriptum szintetizálása az ori H régióban kezdődik, a riboszómális RNS-ek kivágása a hosszú és rövid H-láncú transzkriptumokból történik. A tRNS és az mRNS mindkét DNS-szál transzkriptumainak feldolgozása eredményeként jön létre. A tRNS gének világoszöld színnel vannak feltüntetve.
Szeretné tudni, milyen meglepetéseket hozhat még a mitokondriális genom? Bírság! Olvass tovább!..
A vezető és a 3" nem kódoló régiók, mint a legtöbb nukleáris mRNS. Számos gén tartalmaz intronokat is. Így a citokróm b oxidázt kódoló box génnek két intronja van. Az elsődleges RNS transzkriptumból autokatalitikusan (bármely ill. fehérjék) az első intron nagy részének másolata kivágásra kerül. A megmaradt RNS templátként szolgál a splicingben részt vevő maturáz enzim kialakulásához. Aminosavszekvenciájának egy része az intronok fennmaradó kópiáiban kódolódik. Érettségi hasítások kiiktatják őket, tönkretéve saját mRNS-ét, az exonok másolatai összeolvadnak, és a citokróm-oxidáz b mRNS-e képződik (5. ábra). A jelenség felfedezése arra késztetett bennünket, hogy újragondoljuk az intronokat, mint „semmi kódoló szekvenciákat”.
5. ábra. A citokróm oxidáz b mRNS feldolgozása (érése) élesztő mitokondriumokban. A splicing első szakaszában mRNS képződik, amely szerint a maturáz szintetizálódik, amely a splicing második szakaszához szükséges.
A mitokondriális gének expressziójának vizsgálatakor Trypanosoma brucei meglepő eltérést talált a molekuláris biológia egyik alapvető axiómától, amely azt állítja, hogy az mRNS nukleotidszekvenciája pontosan megfelel a DNS kódoló régióinak szekvenciájának. Kiderült, hogy a citokróm c oxidáz egyik alegységének mRNS-e szerkesztett, azaz a transzkripció után megváltozik az elsődleges szerkezete - négy uracil kerül be. Ennek eredményeként egy új mRNS képződik, amely templátként szolgál az enzim egy további alegységének szintéziséhez, amelynek aminosav-szekvenciájának semmi köze a vírusok, gombák szekvenciájához – vetette össze Burrell angol kutató. az egyik borjú mitokondriális gén szerkezete a citokróm-oxidáz e gén által kódolt alegységében található aminosav-szekvenciával. „ideális”, azaz engedelmeskedik a következő szabálynak: „ha két kodonban két azonos nukleotid van, és a A harmadik nukleotidok ugyanabba az osztályba tartoznak (purin - A, G vagy pirimidin - U, C), akkor ugyanazt az aminosavat kódolják.” Az univerzális kódban két kivétel van e szabály alól: az AUA hármas az izoleucin és az AUG kodon a metionint kódolja, míg az ideális mitokondriális kódban mindkét hármas odyut metionin; az UGG hármas csak triptofánt kódol, míg az UGA hármas stopkodont kódol. Az univerzális kódban mindkét eltérés a fehérjeszintézis alapvető mozzanataihoz kapcsolódik: az AUG kodon beindul, az UGA stopkodon pedig leállítja a polipeptid szintézisét. Az ideális kód nem minden leírt mitokondriumban rejlik, de egyiknek sincs univerzális kódja. Mondhatjuk, hogy a mitokondriumok különböző nyelveket beszélnek, de soha nem a mag nyelvét.
Az "univerzális" genetikai kód és a két mitokondriális kód közötti különbségek
kodon | Mitokondriális emlős kód | Mitokondriális élesztő kód | “ Egyetemes” |
Mint már említettük, a gerinces mitokondriális genom 22 tRNS gént tartalmaz. Hogyan szolgálja ki egy ilyen hiányos készlet az aminosavak mind a 60 kodonját (a 64 triplet ideális kódja négy stopkodont tartalmaz, míg az univerzális kód három)? A tény az, hogy a mitokondriumokban a fehérjeszintézis során a kodon-antikodon kölcsönhatások leegyszerűsödnek - a három antikodon nukleotid közül kettőt használnak a felismeréshez. Így egy tRNS felismeri a kodoncsalád mind a négy képviselőjét, amelyek csak a harmadik nukleotidban különböznek egymástól. Például a GAU antikodont tartalmazó leucin-tRNS a riboszómán a CUU, CUU, CUA és CUG kodonokkal szemben áll, biztosítva a leucin összetéveszthetetlen beépülését a polipeptidláncba. A másik két leucin kodont, az UUA-t és az UUG-t az AAU antikodonnal rendelkező tRNS-ek ismerik fel. Összesen nyolc különböző tRNS-molekula nyolc, egyenként négy kodonból álló családot ismer fel, és 14 tRNS különböző kodonpárokat ismer fel, amelyek mindegyike egy aminosavat kódol.
Fontos, hogy az aminosavaknak a megfelelő mitokondriális tRNS-ekhez való kapcsolódásáért felelős aminoacil-tRNS szintetáz enzimeket a sejtmagban kódolják, és az endoplazmatikus retikulum riboszómáin szintetizálják. Így gerincesekben a polipeptidek mitokondriális szintézisének összes fehérjekomponense a sejtmagban titkosítva van. Ugyanakkor a mitokondriumban a fehérjeszintézist nem gátolja az eukarióta riboszómák munkáját blokkoló cikloheximid, hanem érzékeny az eritromicin és kloramfenikol nevű antibiotikumokra, amelyek gátolják a baktériumok fehérjeszintézisét. Ez a tény az egyik érv amellett, hogy a mitokondriumok aerob baktériumokból származnak az eukarióta sejtek szimbiotikus képződése során.
A saját genetikai rendszerének jelentősége a mitokondriumok számára
Miért van szükségük a mitokondriumoknak saját genetikai rendszerre, míg más organellumoknak, például a peroxiszómáknak és a lizoszómáknak nincs szüksége? Ez a kérdés egyáltalán nem triviális, hiszen egy külön genetikai rendszer fenntartása költséges a sejt számára, tekintettel a szükséges számú további génre a nukleáris genomban. Ide kell kódolni a riboszómális fehérjéket, aminoacil-tRNS szintetázokat, DNS és RNS polimerázokat, RNS feldolgozó és módosító enzimeket stb. A mitokondriumokból származó vizsgált fehérjék többsége aminosav-szekvenciában különbözik a sejt más részeiből származó társaitól, és ott okkal feltételezhető, hogy ezekben az organellumokban nagyon kevés olyan fehérje található, amely máshol is megtalálható. Ez azt jelenti, hogy a mitokondriumok genetikai rendszerének fenntartásához a nukleáris genomnak több tucat további génnek kell lennie. Ennek a "pazarlásnak" az okai nem tisztázottak, és nem vált be az a remény, hogy a nyomot megtalálják a mitokondriális DNS nukleotidszekvenciájában. Nehéz elképzelni, hogy a mitokondriumban képződő fehérjéket miért kell szükségszerűen ott szintetizálni, és nem a citoszolban.
Általában az energiaszervecskék genetikai rendszerének létezését azzal magyarázzák, hogy az organellumokon belül szintetizált fehérjék egy része túlságosan hidrofób ahhoz, hogy kívülről áthaladjon a mitokondriális membránon. Az ATP-szintetáz komplex tanulmányozása azonban azt mutatta, hogy ez a magyarázat valószínűtlen. Bár az ATP-szintetáz egyes fehérjealegységei az evolúció során erősen konzerváltak, szintézisük helyei megváltoznak. A kloroplasztiszokban számos meglehetősen hidrofil fehérje, köztük a komplex F1-ATPáz részének öt alegysége közül négy képződik az organellum riboszómáin. Éppen ellenkezőleg, a gomba Neurospóraállati sejtekben pedig az ATPáz membrán részének egy nagyon hidrofób komponense (9. alegység) szintetizálódik a citoplazma riboszómáin és csak ezt követően kerül át az organellumba. A funkcionálisan ekvivalens fehérjék alegységeit kódoló gének eltérő lokalizációját a különböző szervezetekben nehéz megmagyarázni bármilyen hipotézissel, amely a mitokondriumok és kloroplasztiszok modern genetikai rendszereinek bizonyos evolúciós előnyeit feltételezi.
A fentiek ismeretében csak azt feltételezni kell, hogy a mitokondriumok genetikai rendszere evolúciós zsákutcát jelent. Az endoszimbiózis hipotézis keretében ez azt jelenti, hogy az endoszimbionta gének gazdaszervezet sejtmag genomjába történő átvitelének folyamata leállt, mielőtt teljesen befejeződött volna.
Citoplazmatikus öröklődés
A citoplazmatikus géntranszfer egyes állatokra, köztük az emberekre is súlyosabb következményekkel jár, mint az élesztőkre. Két összeolvadó haploid élesztősejt azonos méretű, és ugyanannyi mitokondriális DNS-sel járul hozzá a létrejövő zigótához. Így élesztőben a mitokondriális genom mindkét szülőtől öröklődik, akik egyformán hozzájárulnak az utódok génállományához (bár több generáció után különálló a leszármazottak gyakran csak az egyik szülőtípus mitokondriumát tartalmazzák). Ezzel szemben a magasabb rendű állatokban a petesejt több citoplazmát ad a zigótához, mint a spermium, és egyes állatokban a spermium egyáltalán nem járul hozzá a citoplazmához. Ezért azt gondolhatjuk, hogy magasabbrendű állatokban a mitokondriális genom csak az egyik szülőtől kerül átvitelre (nevezetesen anyai vonalak); valóban, ezt kísérletileg is megerősítették. Kiderült például, hogy ha két laboratóriumi vonal patkányait keresztezzük a nukleotidszekvenciában kissé eltérő mitokondriális DNS-sel (A és B típus), akkor olyan utódokat kapunk, amelyek
csak anyai típusú mitokondriális DNS.
A citoplazmatikus öröklődés, ellentétben a sejtmaggal, nem engedelmeskedik Mendel törvényeinek. Ez annak a ténynek köszönhető, hogy a magasabbrendű állatokban és növényekben a különböző nemű ivarsejtek eltérő mennyiségű mitokondriumot tartalmaznak. Tehát az egér tojásában 90 ezer mitokondrium van, a spermában pedig csak négy. Nyilvánvaló, hogy a megtermékenyített petesejtben a mitokondriumok túlnyomórészt vagy csak nőstényből származnak, azaz minden mitokondriális gén anyai öröklődésű. A citoplazmatikus öröklődés genetikai elemzése nehéz a sejtmag-citoplazma kölcsönhatások miatt. Citoplazmatikus hímsterilitás esetén a mutáns mitokondriális genom kölcsönhatásba lép bizonyos nukleáris génekkel, amelyek recesszív alléljai szükségesek a tulajdonság kialakulásához. E gének domináns alléljai mind homo-, mind heterozigóta állapotban visszaállítják a növények termékenységét, függetlenül a mitokondriális genom állapotától.
Egy konkrét példával szeretnék kitérni a gének anyai öröklődésének mechanizmusára. Annak érdekében, hogy véglegesen és visszavonhatatlanul megértsük a mitokondriális gének nem mendeli (citoplazmatikus) öröklődésének mechanizmusát, gondoljuk át, mi történik az ilyen génekkel, amikor két haploid sejt összeolvad, és diploid zigótát alkot. Abban az esetben, ha az egyik élesztősejt olyan mutációt hordoz, amely meghatározza a mitokondriális fehérjeszintézis kloramfenikollal szembeni rezisztenciáját, a másik, egy vad típusú sejt pedig erre az antibiotikumra érzékeny: a mutáns gének könnyen azonosíthatók, ha élesztőt tenyésztéssel hordozó táptalajon. glicerin, amelyet csak ép mitokondriumokkal rendelkező sejtek használhatnak; ezért kloramfenikol jelenlétében csak a mutáns gént hordozó sejtek tudnak ilyen táptalajon növekedni. A diploid zigótánk kezdetben mutáns és vad típusú mitokondriumokkal is rendelkezik. A mitózis hatására a zigótából egy diploid leánysejt rügyez ki, amely csak kevés mitokondriumot tartalmaz majd. Több mitotikus ciklus után végül az egyik új sejt megkapja az összes mitokondriumot, akár mutáns, akár vad típusú. Ezért egy ilyen sejt minden utódjának genetikailag azonos mitokondriuma lesz. Egy ilyen véletlenszerű folyamatot, amelynek eredményeként csak egyféle mitokondriumot tartalmazó diploid utódok képződnek, az ún. mitotikusth segyülekezésth. Amikor egy diploid sejtben csak egyfajta mitokondrium van, meiózison megy keresztül, mind a négy haploid leánysejt ugyanazokat a mitokondriális géneket kapja. Az öröklődésnek ezt a fajtáját ún nemendeegy oroszlán sovány vagy citoplazmatikus ellentétben a nukleáris gének mendeli öröklődésével. A citoplazmatikus típus szerinti gének átvitele azt jelenti, hogy a vizsgált gének a mitokondriumban helyezkednek el.
Az eukarióta sejt és a szervezet egészének bonyolult hierarchikus szerveződésének megértéséhez szükséges a mitokondriális genomok, evolúciójuk, a populációgenetika sajátos törvényszerűségei, a nukleáris és a mitokondriális genetikai rendszerek kapcsolatának vizsgálata.
Egyes örökletes betegségek és az emberi öregedés összefüggésbe hozható a mitokondriális DNS vagy a mitokondriumok működését szabályozó nukleáris gének bizonyos mutációival. Gyűlnek az adatok az mtDNS-hibáknak a karcinogenezisben való részvételéről. Ezért a mitokondriumok a rák kemoterápia célpontjai lehetnek. Vannak tények a nukleáris és mitokondriális genomok szoros kölcsönhatásáról számos emberi patológia kialakulásában. Súlyos izomgyengeségben, ataxiában, süketségben, mentális retardációban szenvedő, autoszomális domináns módon öröklődő betegeknél többszörös mtDNS deléciót találtak. Szexuális dimorfizmust állapítottak meg a szívkoszorúér-betegség klinikai megnyilvánulásaiban, ami nagy valószínűséggel az anyai hatásnak – a citoplazmatikus öröklődésnek – köszönhető. A génterápia fejlődése reményt ad a mitokondriális genom hibáinak belátható időn belüli korrekciójára.
Mint ismeretes, a többkomponensű rendszer egyik összetevőjének működésének ellenőrzéséhez szükségessé válik ennek az összetevőnek a megszüntetése a bekövetkezett változások utólagos elemzésével. Mivel jelen esszé témája az anyai genom szerepének jelzése az utódok fejlődésében, logikus lenne megismerni a mitokondriális genom összetételében bekövetkező különböző tényezők által okozott zavarok következményeit. A mutációs folyamat a fenti szerep tanulmányozásának eszközévé vált, és működésének számunkra érdekes következményei az ún. mitokondriális betegségek.
A mitokondriális betegségek az emberi citoplazmatikus öröklődés, vagy inkább "organellum öröklődés" példái. Ezt a pontosítást meg kell tenni, mert mostanra bebizonyosodott, hogy – legalábbis egyes organizmusokban – léteznek olyan citoplazmatikus örökletes determinánsok, amelyek nem kapcsolódnak a sejtszervecskékhez - citogének (-Vechtomov, 1996).
A mitokondriális betegségek a betegségek heterogén csoportja, amelyet a mitokondriumok genetikai, szerkezeti, biokémiai hibái és a szöveti légzés zavara okoznak. Az átfogó genealógiai, klinikai, biokémiai, morfológiai és genetikai elemzés fontos a mitokondriális betegség diagnózisának felállításához. A mitokondriális patológia fő biokémiai jele a tejsavas acidózis kialakulása, általában hiperlaktát-acidémiát észlelnek hiperpiruvát-acidémiával kombinálva. A különböző változatok száma elérte a 120 formát. Az agy-gerincvelői folyadékban a tejsav és a piroszőlősav koncentrációja stabilan növekszik.
A mitokondriális betegségek (MD) jelentős problémát jelentenek a modern orvostudomány számára. Az örökletes átvitel módszerei szerint az MB-k között vannak olyan mendeli típus szerint monogénen öröklődő betegségek, amelyekben a nukleáris gének mutációja miatt a mitokondriális fehérjék szerkezete, működése vagy megzavarodik, vagy a mitokondriális expressziója. DNS-elváltozások, valamint a mitokondriális gének mutációi által okozott betegségek, amelyek többnyire az anyai vonalon keresztül szállnak át az utódokra.
A morfológiai vizsgálatok adatai a mitokondriumok durva patológiájára utalnak: abnormális mitokondriumok proliferációja, a mitokondriumok alakjának és méretének megsértésével járó polimorfizmus, a kristályok dezorganizációja, abnormális mitokondriumok felhalmozódása a szarkolemma alatt, parakristályos zárványok, mitokondriumok közötti zárványok vakuolák
A mitokondriális betegségek formái
1 . A mitokondriális DNS mutációi által okozott mitokondriális betegségek
1.1 A mitokondriális DNS deléciói által okozott betegségek
1.1.1.Kearns-Sayre szindróma
A betegség 4-18 éves korban jelentkezik, progresszív külső ophthalmoplegia, retinitis pigmentosa, ataxia, szándékos tremor, atrioventricularis szívblokk, a liquor 1 g/l-nél nagyobb fehérjeszintje, "rongyos" vörös rostok a vázban. izombiopsziák
1.1.2 Pearson-szindróma
A betegség debütálása a születéstől vagy az élet első hónapjaiban, néha előfordulhat encephalomyopathiák, ataxia, demencia, progresszív külső ophthalmoplegia, hypoplasiás vérszegénység, károsodott exokrin hasnyálmirigy-funkció, progresszív lefolyás
2 .A mitokondriális DNS pontmutációi által okozott betegségek
Anyai öröklődés, akut vagy szubakut látásélesség csökkenés az egyik vagy mindkét szemen, neurológiai és osteoartikuláris rendellenességekkel kombinálva, retina mikroangiopátia, progresszív lefolyás a remisszió vagy a látásélesség helyreállításának lehetőségével, a betegség kezdete 20 éves korban -30 év
2.2. NAPR-szindróma (neuropathia, ataxia, retinitis pigmentosa)
Anyai öröklődés, neuropátia, ataxia és retinitis pigmentosa kombinációja, pszichomotoros retardáció, demencia, "szakadt" vörös rostok jelenléte az izomszövet biopsziában
2.3 MERRF szindróma (mioklónus-epilepszia, "szakadt" vörös rostok)
Anyai öröklődés típusa, a betegség 3-65 éves korban kezdete, myoclonus epilepszia, ataxia, demencia neuroszenzoros süketséggel kombinálva, látóidegsorvadás és mélyérzékenységi zavarok, tejsavas acidózis, EEG vizsgálattal generalizált fürdő epilepsziás komplexek derülnek ki , "szakadt" vörös rostok vázizom biopsziás mintákban, progresszív lefolyás
2.4. MELAS-szindróma (mitokondriális encephalomyopathia, tejsavas acidózis, stroke-szerű epizódok)
Anyai típusú öröklődés, a betegség 40 éves kor előtti kezdete, terhelési intolerancia, migrénszerű fejfájás hányingerrel, hányással, stroke-szerű epizódok, görcsök, tejsavas acidózis, "rongyos" vörös rostok az izombiopsziákban, progresszív lefolyás.
3 .Az intergenomikus kommunikáció hibáihoz kapcsolódó patológia
3.1 A mitokondriális DNS többszörös deléciójának szindrómái
Blepharoptosis, külső ophthalmoplegia, izomgyengeség, szenzorineurális süketség, látóideg atrófia, progresszív lefolyás, "szakadt" vörös rostok a vázizom biopsziában, a légzési lánc enzimek aktivitásának csökkenése.
3.2 Mitokondriális DNS-deléciós szindróma
Autoszomális recesszív öröklődési minta
Klinikai formák:
3.2.1.végzetes infantilis
a) súlyos májelégtelenség b) hepatopathia c) izom-hipotenzió
Debütálás az újszülöttkori időszakban
3.2.2.veleszületett myopathia
Súlyos izomgyengeség, generalizált hipotenzió, kardiomiopátia és görcsök, vesekárosodás, glucosuria, aminoacidopathia, phosphaturia
3.2.3.Infantilis myopathia
az első 2 életévben jelentkezik, progresszív izomgyengeség, proximális izomcsoportok sorvadása és az ínreflexek elvesztése, gyorsan progresszív lefolyás, halál az első 3 életévben.
4 .Magi DNS-mutációk által okozott mitokondriális betegségek
4.1 A légzési lánc hibáival kapcsolatos betegségek
4.1.1. Az 1. komplex hiánya (NADH:CoQ-reduktáz)
15 éves kor előtti megjelenés, myopathia szindróma, pszichomotoros retardáció, szív- és érrendszeri betegségek, terápiarezisztens rohamok, többszörös neurológiai rendellenességek, progresszív lefolyás
4.1.2 A 2-es komplex hiánya (szukcinát-CoQ-reduktáz)
Az encephalomyopathia szindróma jellemzi, progresszív lefolyás, görcsök, ptosis alakulhat ki
4.1.3 A 3-as komplex hiánya (CoQ-citokróm C-oxidoreduktáz)
Multiszisztémás rendellenességek, különböző szervek és rendszerek károsodása, a központi és perifériás idegrendszer, az endokrin rendszer, a vesék érintettségével, progresszív lefolyás
4.1.4. A komplex hiánya (citokróm C-oxidáz)
4.1.4.1 Halálos csecsemőkori veleszületett tejsavas acidózis
Mitokondriális myopathia veseelégtelenséggel vagy kardiomiopátiával, újszülött korban kezdődik, súlyos légzési rendellenességek, diffúz izom hipotenzió, progresszív lefolyás, halál az első életévben.
4.1.4.2.Jóindulatú infantilis izomgyengeség
Az atrófia megfelelő és időben történő kezeléssel gyorsan stabilizálja a folyamatot és 1-3 életévre felépül.
5 Menkes szindróma (trichopolyodystrophia)
A pszichomotoros fejlődés éles késése, növekedési retardáció, disztrófiás növekedés és hajelváltozások,
6 . Mitokondriális encephalomyopathiák
6.1.Leigh szindróma(szubakut neurotikus encephalomyelopathia)
A 6 hónapos élet után manifesztálódó izom hipotenzió, ataxia, nystagmus, piramis tünetek, ophthalmoplegia, látóideg-sorvadás, kardiomiopátia és enyhe metabolikus acidózis.
6.2.Alpers szindróma(progresszív szklerotizáló polidystrophia)
Az agy szürkeállományának degenerációja májcirrózissal kombinálva, az 5-ös komplex (ATP-szintetáz) hiánya, pszichomotoros retardáció, ataxia, demencia, izomgyengeség, a betegség progresszív lefolyása, kedvezőtlen prognózis
6.3 Koenzim-Q hiány
Anyagcsere-krízisek, izomgyengeség és -fáradtság, ophthalmoplegia, süketség, látásromlás, stroke-szerű epizódok, ataxia, myoclonus epilepszia, vesekárosodás: glucosuria, aminoacidopathia, phosphaturia, endokrin betegségek, progresszív lefolyás, csökkent légzési lánc enzimaktivitás
7 .A tej- és piroszőlősav károsodott metabolizmusával kapcsolatos betegségek
7.1 Piruvát-karboxiláz hiánya Autoszomális recesszív típusú öröklődés, a betegség újszülöttkori megjelenése, "lomha gyermek" tünetegyüttese, terápiának ellenálló görcsök, magas ketontestek koncentrációja a vérben, hiperammonémia, hiperlizinémia, csökkent piruvát-karboxiláz aktivitása a vázizmokban
7.2 Piruvát-dehidrogenáz hiány
Újszülöttkori megnyilvánulások, craniofacialis dysmorphia, terápiának ellenálló görcsök, légzési és szopási zavarok, "lomha gyermek" tünetegyüttes, agyi diszgenezis, súlyos acidózis magas laktát és piruvát tartalommal
7.3 A piruvát-dehidrogenáz aktivitásának csökkenése
Az első életévben kialakuló mikrokefália, pszichomotoros retardáció, ataxia, izomdystonia, choreoathetosis, magas piruvát tejsavas acidózis
7.4 Dihidrolipoil-transzacetiláz hiány
Autoszomális recesszív öröklődés, a betegség kezdete az újszülött korban, mikrokefália, pszichomotoros retardáció, izom hipotenzió az izomtónus növekedésével, a porckorongok sorvadása, tejsavas acidózis, a dihidrolipoil-transzacetiláz aktivitásának csökkenése
7.5 Dihidrolipoil-dehidrogenáz hiány
Autoszomális recesszív típusú öröklődés, a betegség kezdete az első életévben, "lomha gyermek" tünetegyüttes, hányással és hasmenéssel járó dysmetaboliás krízisek, pszichomotoros retardáció, porckorongok sorvadása, tejsavas acidózis, emelkedett alaninszint a vérben szérum, α-ketoglutarát, elágazó láncú α-keto savak, csökkent dihidrolipoil-dehidrogenáz aktivitás
8 .A zsírsavak béta-oxidációjának hibáiból eredő betegségek
8.1 Hosszú láncú acetil-CoA dehidrogenáz hiánya
Autoszomális recesszív típusú öröklődés, a betegség kezdete az élet első hónapjaiban, hányással és hasmenéssel járó anyagcsere-válságok, "lomha gyermek" tünetegyüttes, hipoglikémia, dikarbonsav-aciduria, a hosszú szénláncú zsírsav-acetil-CoA dehidrogenáz aktivitásának csökkenése
8.2 Közepes láncú acetil-CoA dehidrogenáz hiánya
Autoszomális recesszív típusú öröklődés, a betegség újszülött korban vagy az élet első hónapjaiban kialakuló, hányással és hasmenéssel járó anyagcsere-válságok,
izomgyengeség és hipotónia, gyakran alakul ki hirtelen halál szindróma, hipoglikémia, dikarbonsav aciduria, közepes szénláncú zsírsav-acetil-CoA dehidrogenáz aktivitás csökkenése
8.3. Rövid szénláncú zsírsav-acetil-CoA dehidrogenáz hiány
Autoszomális recesszív öröklődés, a betegség kezdetének eltérő kora, csökkent terhelési tolerancia, hányással és hasmenéssel járó anyagcsere-krízisek, izomgyengeség és hipotenzió, a metil-borostyánkősav fokozott vizeletürítése, rövid szénláncú zsírsavak acetil-CoA-dehidrogenáza
8.4 Az acetil-CoA zsírsav-dehidrogenázok többszörös hiánya
újszülött forma: craniofacialis dysmorphia, agyi dysgenesis, súlyos hipoglikémia és acidózis, rosszindulatú lefolyás, a zsírsavak összes acetil-CoA dehidrogenázának csökkent aktivitása,
Infantilis forma:"lomha gyerek" tünetegyüttes, kardiomiopátia, anyagcsere-válságok, hipoglikémia és acidózis
8.5 Az összes zsírsav-acetil-CoA-dehidrogenáz aktivitásának csökkenése
Késői debütáló forma: az izomgyengeség időszakos epizódjai, az anyagcsere-válságok, a hipoglikémia és az acidózis kevésbé kifejezettek, az intelligencia megmarad,
9 .A Krebs-ciklus fermentopátiái
9.1 Fumaráz-hiány
Autoszomális recesszív öröklődés, újszülöttkori vagy újszülöttkori megjelenés, mikrokefália, generalizált izomgyengeség és hipotenzió, letargia epizódok, gyorsan progresszív encephalopathia, rossz prognózis
9.2 A szukcinát-dehidrogenáz hiánya
Ritka betegség, amelyet progresszív encephalomyopathia jellemez
9.3 Alfa-ketoglutarát-dehidrogenáz hiány
Autoszomális recesszív típusú öröklődés, a betegség újszülöttkori debütálása, mikrokefália, "lomha baba" tünetegyüttes, letargia epizódok, tejsavas acidózis, gyorsan progresszív lefolyás, csökkent Krebs-ciklus enzimtartalom a szövetekben
9.4 A karnitin és az anyagcsere enzimek hiányának szindrómái
Karnitin-palmitoiltranszferáz-1 hiány, autoszomális recesszív öröklődés, a betegség korai kezdete, nem ketonémiás hipoglikémiás kóma, hepatomegalia, hipertrigliceridémia és mérsékelt hyperammonemia, a karnitin-palmitoiltranszferáz-1 aktivitásának csökkenése a fibroblasztokban és a májsejtekben
9.5 A karnitin acilkarnitin transzlokáz hiánya
A betegség korai megjelenése, szív- és érrendszeri és légúti rendellenességek, "lomha gyermek" tünetegyüttes, letargia és kóma epizódok, megnövekedett karnitin-észter-koncentráció és hosszú szénlánc a szabad karnitin csökkenése hátterében a vérszérumban, csökkent aktivitás karnitin-acilkarnitin transzlokáz
9.6 A karnitin-palmitoiltranszferáz-2 hiánya
Autoszomális recesszív öröklődés, izomgyengeség, myalgia, myoglobinuria, a karnitin-palmitoil-transzferáz-2 aktivitásának csökkenése a vázizmokban
Autoszomális recesszív öröklődés, myopathiás tünetegyüttes, letargia és letargia epizódok, kardiomiopátia, hipoglikémia epizódok, csökkent szérum karnitinszint és fokozott vizeletkiválasztás.
A mitokondriális (és nem csak) genom működésének bizonyos változásaihoz kapcsolódó patológiák ilyen „szörnyű” listájának elemzése után bizonyos kérdések merülnek fel. Mik a mitokondriális gének termékei, és milyen szuper-mega-életfontosságú sejtfolyamatokban vesznek részt?
Mint kiderült, a fenti patológiák némelyike a NADH-dehidrogenáz komplex 7 alegységének, az ATP szintetáz 2 alegységének, a citokróm c oxidáz 3 alegységének és az ubikinol-citokróm c-reduktáz (citokrometáz) 1 alegységének a szintézisének károsodása esetén fordulhat elő. ), amelyek a mitokondriumok géntermékei. Ennek alapján megállapíthatjuk, hogy ezek a fehérjék kulcsszerepet játszanak a sejtlégzés, a zsírsavoxidáció és az ATP szintézis folyamataiban, az elektrontranszfer a belső mt membrán elektrontranszport rendszerében, az antioxidáns rendszer működésében stb.
Az apoptózis mechanizmusaira vonatkozó legfrissebb adatok alapján sok tudós arra a következtetésre jutott, hogy az apoptózis irányító központja pontosan ...
A mitokondriális fehérjék szerepét az MT szintézist gátló antibiotikumok alkalmazásában is kimutatták. Ha a szövettenyészetben lévő emberi sejteket antibiotikummal, például tetraciklinnel vagy kloramfenikollal kezelik, akkor egy-két osztódás után növekedésük leáll. Ennek oka a mitokondriális fehérjeszintézis gátlása, ami hibás mitokondriumok megjelenéséhez és ennek következtében az ATP elégtelen képződéséhez vezet. Akkor miért használhatók az antibiotikumok a bakteriális fertőzések kezelésére? Erre a kérdésre több válasz is létezik:
1. Egyes antibiotikumok (például eritromicin) nem jutnak át az emlős mitokondriumok belső membránján.
2. Testünk sejtjeinek nagy része nem, vagy nagyon lassan osztódik, így a meglévő mitokondriumok újakra cserélődése is ugyanolyan lassú (sok szövetben a mitokondriumok fele körülbelül öt nap alatt vagy még tovább cserélődik). Így a normál mitokondriumok száma csak akkor fog kritikus szintre csökkenni, ha a mitokondriális fehérjeszintézis blokádja több napig fennmarad.
3. A szöveten belül bizonyos körülmények megakadályozzák bizonyos gyógyszerek behatolását a legérzékenyebb sejtek mitokondriumaiba. Például a csontvelőben a Ca2+ magas koncentrációja Ca2+-tetraciklin komplex képződéséhez vezet, amely nem tud behatolni a gyorsan osztódó (és ezért a legsebezhetőbb) vérsejtek prekurzoraiba.
Ezek a tényezők lehetővé teszik bizonyos, a mitokondriális fehérjeszintézist gátló gyógyszerek antibiotikumként való alkalmazását magasabb rendű állatok kezelésében. Ezen gyógyszerek közül csak kettőnek van mellékhatása: a nagy dózisú kloramfenikollal végzett hosszú távú kezelés a csontvelő hematopoietikus működésének károsodásához vezethet (elnyomja a vörösvértestek és fehérvérsejtek képződését), valamint a tetraciklin hosszú távú alkalmazása. károsíthatja a bélhámot. De mindkét esetben még mindig nem teljesen világos, hogy ezeket a mellékhatásokat a mitokondriális biogenezis blokádja vagy más okok okozzák.
Következtetés
Az mt genom szerkezeti és funkcionális jellemzői a következők. Először is megállapították, hogy az mtDNS-t az anyától mindenkihez továbbítják
leszármazottaitól és lányaitól az összes következő nemzedékre, de a fiúk nem adják tovább DNS-üket (anyai örökség). anyai jellem
Az mtDNS öröklődés valószínűleg két körülményre vezethető vissza: vagy olyan kicsi az apai mtDNS aránya (nem
25 ezer anyai mtDNS-enként egynél több DNS-molekula), hogy meglévő módszerekkel nem mutathatók ki, vagy a megtermékenyítés után az apai mitokondriumok replikációja blokkolva van. Másodszor, a kombinatív variabilitás hiánya - az mtDNS csak az egyik szülőhöz tartozik, ezért a meiózisban nincsenek a nukleáris DNS-re jellemző rekombinációs események, és a nukleotidszekvencia nemzedékről nemzedékre változik csak a mutációk miatt. Harmadszor, az mtDNS-nek nincsenek intronjai.
(nagy a valószínűsége annak, hogy véletlenszerű mutáció éri a DNS kódoló régióját), védő hisztonok és hatékony DNS-javító rendszer – mindez 10-szer nagyobb mutációs rátát határoz meg, mint a nukleáris DNS-ben. Negyedszer, a normál és a mutáns mtDNS egyidejűleg létezhet egy sejten belül – ez a heteroplazmia jelensége (a csak normális vagy csak mutáns mtDNS jelenlétét homoplazmiának nevezzük). Végül mindkét lánc átíródik és mtDNS-vé alakul, és az mtDNS genetikai kódja számos jellemzőben eltér az univerzálistól (az UGA triptofánt, az AUA a metionint kódolja, az AGA és az AGG pedig stop-
kodonok).
Az mt genom ezen tulajdonságai és fent említett funkciói az mtDNS nukleotid szekvencia variabilitásának tanulmányozását felbecsülhetetlen eszközzé tették az orvosok, törvényszéki tudósok, evolúciós biológusok,
a történettudomány képviselőit konkrét feladataik megoldásában.
1988 óta, amikor felfedezték, hogy az mtDNS génmutációk állnak a mitokondriális myopathiák (JY Holt és mtsai, 1988) és a Leber-féle örökletes optikai neuropátia (DC Wallace, 1988) hátterében, a humán mt genom mutációinak további szisztematikus azonosítása az emberi mt genomban a mutációk kialakulásához vezetett. a mitokondriális betegségek (MD) fogalma. Jelenleg patológiás mtDNS mutációkat fedeztek fel minden típusú mitokondriális génben.
Bibliográfia
1. Skulachev, mitokondriumok és oxigén, Soros. oktat. magazin
2. A biokémia alapjai: Három kötetben, M.: Mir, .
3. Nicholes D. G. Bioenergetics, Bevezetés. a kemiozmushoz. Th., Acad. Nyomda, 1982.
4. Stryer L. Biochemistry, 2. kiadás. San Francisco, Freeman, 1981.
5. Skulachev biológiai membránok. M., 1989.
6., Csencov retikulum: Szerkezet és néhány funkció // Tudomány eredményei. A biológia általános problémái. 1989
7. Csencov citológia. M.: Moszkvai Állami Egyetem Kiadója, 1995
8. , A mitokondriális genom illetékességi köre // Vestn. RAMN, 2001. ‹ 10. S. 31-43.
9. Holt I. J, Harding A. E., Morgan-Hughes I. A. Az izom-mitokondriális DNS törlése mitokondriális myopathiában szenvedő betegeknél. Nature, 1988, 331:717-719.
10. satöbbi. Az emberi genom és a hajlam gének. SPb., 2000
11. , Mitokondriális genom. Novoszibirszk, 1990.
12. // Soros. oktat. magazin 1999. 10. sz. pp.11-17.
13. A szimbiózis szerepe a sejtevolúcióban. M., 1983.
14. // Soros. oktat. magazin 1998. 8. sz. S.2-7.
15. // Soros. oktat. magazin 2000. 1. sz. S.32-36.
Kijevi Nemzeti Egyetem. Tarasz Sevcsenko
Biológiai Tanszék
absztrakt
a témában:
„Az anyai genom szerepe az utódok fejlődésében”
Val velottenta IVtanfolyam
Biokémiai Tanszék
Frolova Artem
Kijev 2004
Terv:
Bevezetés................................................. ..............................egy
A mitokondriumok eredetének szimbiotikus elmélete ...... 2
A sejtmag szerepe a mitokondriális biogenezisben ................................................ ......................5
A mitokondriumok szállítási rendszerei .................................................. ..............................7
A mitokondriális genomok mérete és alakja ..................................10
A mitokondriális genom működése...................14
A saját genetikai rendszer jelenlétének jelentősége a mitokondriumok számára ................................................ ...................................................... .......tizenkilenc
Citoplazmatikus öröklődés................................................20
Történelmileg az első ilyen jellegű vizsgálatot mitokondriális DNS felhasználásával végeztek. A tudósok Afrika, Ázsia, Európa, Amerika őslakosaitól vettek mintát, és ebben a kezdetben kis mintában a különböző egyedek mitokondriális DNS-ét hasonlították össze egymással. Azt találták, hogy Afrikában a legmagasabb a mitokondriális DNS-diverzitás. És mivel ismert, hogy a mutációs események megváltoztathatják a mitokondriális DNS típusát, és az is ismert, hogyan változhat, ezért meg lehet mondani, hogy melyik embertípus melyikből mutálódhat. A DNS-teszteken átesett emberek közül az afrikaiak találtak sokkal nagyobb eltérést. Más kontinenseken a mitokondriális DNS-típusok kevésbé változatosak voltak. Ez azt jelenti, hogy az afrikaiaknak több idejük volt felhalmozni ezeket a változásokat. Több idejük volt a biológiai evolúcióra, ha éppen Afrikában találnak olyan ősi DNS-maradványokat, amelyek nem jellemzőek az európai emberi mutációkra.
Vitatható, hogy a mitokondriális DNS-t használó genetikusoknak sikerült bizonyítaniuk a nők afrikai származását. Az Y kromoszómákat is tanulmányozták. Kiderült, hogy Afrikából is érkeznek férfiak.
A mitokondriális DNS-vizsgálatoknak köszönhetően nemcsak azt lehet megállapítani, hogy egy személy Afrikából származott, hanem az is, hogy mikor jött létre. Az emberiség mitokondriális ősanyja megjelenésének idejét a csimpánzok és a modern ember mitokondriális DNS-ének összehasonlító vizsgálata alapján határozták meg. Ismerve a mutációs eltérés mértékét - 2-4% millió évenként - meg lehet határozni a két ág, a csimpánz és a mai ember elválasztásának idejét. Ez körülbelül 5-7 millió évvel ezelőtt történt. Feltételezzük, hogy a mutációs divergencia sebessége állandó.
Mitokondriális Éva
Amikor az emberek mitokondriális Éváról beszélnek, nem az egyénre gondolnak. Arról beszélnek, hogy az evolúció során hasonló tulajdonságokkal rendelkező egyedek egész populációja jött létre. Úgy gondolják, hogy a mitokondriális Éva olyan időszakban élt, amikor őseink száma hirtelen, körülbelül tízezer egyedre csökkent.
A fajok eredete
A különböző populációk mitokondriális DNS-ének tanulmányozása során a genetikusok azt javasolták, hogy az ősi populációt még Afrika elhagyása előtt három csoportra osztották, ami három modern fajt - afrikai, kaukázusi és mongoloid - eredményezett. Úgy gondolják, hogy ez körülbelül 60-70 ezer évvel ezelőtt történt.
A neandervölgyiek és a modern emberek mitokondriális DNS-ének összehasonlítása
Az ember eredetével kapcsolatban további információkhoz jutottak a neandervölgyi és a modern ember mitokondriális DNS-ének genetikai szövegeinek összehasonlításával. A tudósoknak sikerült elolvasniuk két neandervölgyi ember csontmaradványának mitokondriális DNS-ének genetikai szövegét. Az első neandervölgyi ember csontjait a németországi Feldhover-barlangban találták meg. Kicsit később egy neandervölgyi gyermek mitokondriális DNS-ének genetikai szövegét olvasták fel, amelyet az Észak-Kaukázusban, a Mezhmayskaya barlangban találtak. A modern emberek és a neandervölgyiek mitokondriális DNS-ének összehasonlításakor nagyon nagy különbségeket találtak. Ha egy DNS-szakaszt veszünk, akkor a 370 nukleotidból 27 különbözik. És ha összehasonlítjuk egy modern ember genetikai szövegét, mitokondriális DNS-ét, akkor mindössze nyolc nukleotid különbözik. Úgy tartják, hogy a neandervölgyi és a modern ember teljesen külön ág, mindegyik fejlődése független volt egymástól.
A neandervölgyi és a modern ember mitokondriális DNS-ének genetikai szövegeinek különbségének tanulmányozása során megállapították e két ág szétválásának időpontját. Ez körülbelül 500 ezer évvel ezelőtt történt, és körülbelül 300 ezer éve történt végső szétválásuk. Úgy tartják, hogy a neandervölgyiek Európában és Ázsiában telepedtek le, és kiszorították őket a modern emberek, akik 200 ezer évvel később hagyták el Afrikát. És végül, körülbelül 28-35 ezer évvel ezelőtt, a neandervölgyiek kihaltak. Hogy miért történt ez, általában még nem világos. Talán nem bírták ki a versenyt egy modern típusú emberrel, vagy ennek más okai voltak.
A fogyasztás ökológiája. Egészség: Haplocsoport - hasonló haplotípusok csoportja, amelyeknek közös őse van, és amelyben ugyanaz a mutáció ment végbe mindkét haplotípusban ...
Gyerekkoromban megkérdeztem a nagymamát a gyökerekről, ő mesélt egy legendát, miszerint távoli dédnagyapja egy "helyi" lányt vett feleségül. Érdekelt ez a dolog, és egy kis kutatásba kezdtem. A Vologda régióban élő vepszei finnugorok. A családi legenda pontos tesztelése érdekében a genetikához fordultam. És megerősítette a családi legendát.
Haplogroup (az emberi populáció genetikájában - az emberiség genetikai történetét tanulmányozó tudomány) - hasonló haplotípusok csoportja, amelyek közös őssel rendelkeznek, amelyben mindkét haplotípusban ugyanaz a mutáció ment végbe. A "haplocsoport" kifejezést széles körben használják a genetikai genealógiában, ahol az Y-kromoszómális (Y-DNS), mitokondriális (mtDNS) és MHC haplocsoportokat tanulmányozzák. Az Y-DNS genetikai markerei az Y-kromoszómával kizárólag az apai vonalon (azaz az apától a fiúkig), az mtDNS-markerek pedig az anyai vonalon keresztül (az anyától minden gyermekhez) továbbadódnak.
A mitokondriális DNS (mtDNS) anyáról gyermekre kerül. Mivel csak a nőstények tudják átadni az mtDNS-t utódaiknak, az mtDNS-teszt a közvetlen anyai vonalon keresztül információt ad az anyáról, az anyjáról stb. Férfiak és nők is kapnak mtDNS-t az anyjuktól, így férfiak és nők is részt vehetnek az mtDNS tesztelésében. Bár mutációk előfordulnak az mtDNS-ben, gyakoriságuk viszonylag alacsony. Az évezredek során ezek a mutációk felhalmozódtak, és emiatt az egyik családban a női vonal genetikailag különbözik a másiktól. Miután az emberiség megtelepedett a bolygón, a mutációk továbbra is véletlenszerűen jelentek meg az egykor egyetlen emberi faj egymástól távol eső populációiban.
A mitokondriális haplocsoportok migrációja.
orosz észak.
Nagyon közel áll hozzám az orosz észak történelme, természete és kultúrája. Ez annak is köszönhető, hogy onnan származik a nagymamám, aki velünk élt, és sok időt szentelt a nevelésemnek. De azt gondolom, hogy a fehéroroszok számára a közelség még nagyobb: végül is az orosz északi részét a Krivicsek lakták, akik egyben a leendő Fehéroroszország magját is alkották. Ráadásul Pszkov és Novgorod ősi szláv központok, bizonyos mértékig demokratikusak, saját vecsével (akárcsak Kijev és Polock).
Elég, ha felidézzük a Pszkov Vecse Köztársaság és a Novgorodi Köztársaság történetét. Ezek a területek sokáig a Litván Nagyhercegség és a Moszkvai Hercegség között ingadoztak, de az utóbbi ragadta meg a kezdeményezést a "földgyűjtésben". Más körülmények között ennek a régiónak az identitása önálló nemzetiséggé fejlődhet. Sokan azonban büszkén nevezik magukat "északi oroszoknak". Néhány fehéroroszhoz hasonlóan megkülönböztetik Nyugat-Belorusz (Litvánia, Litvinek) Kelet-Belorussziát (ruszinok). Kérem, ne keressen politikai hátteret szavaimban.
Ha Fehéroroszországban a szlávok keveredtek a balti törzsekkel, akkor Oroszországban - a finnugorokkal. Ez biztosította a különböző régiók egyedi etnikai hovatartozását. A szomszédos falvakból származó Parfjonov nagyon pontosan mondta: „Mindig érzem a származásomat. Észak-orosz - számomra nagyon fontos. Ez az én elképzelésem Oroszországról, a jellemünkről, az etikáról és az esztétikáról. Voronyezstől délre számomra más oroszok. Kíváncsi vagyok, hogy vannak Parfenovok a családomban. Aksinya Parfenova (1800-1904) Kirill Kirillovics Koricsev (Aleksandra Alekszejevna Zemskova férje) nagymamája. Ez a vezetéknév azonban gyakori, így talán rokonok, vagy talán nem.
Cserepovec, balra a dédnagymama, jobbra lent a nagymama, 1957?
A mitokondriális csoportom a D5a3a.
A GVS1 szekvenálásánál - 16126s, 16136s, 16182s, 16183s, 16189s, 16223T, 16360T, 16362S. Ez azt jelenti, hogy a mitokondriális csoportom a D5a3a. Ez egy nagyon ritka haplocsoport, még a genetikusok is meglepődtek – ez az első alkalom, hogy ilyen csoportot határoztak meg Fehéroroszországban. Általában D egy ázsiai csoport. A tudósok azt írják, hogy Észak-Eurázsia csak néhány etnikai csoportjának génállományában található meg.
Egyetlen D5a3 vonalat találtak a tádzsikoknál, az altájiaknál, a koreaieknél és a nagyszabású Novgorod-i oroszoknál. Mindegyikre (a koreai kivételével) jellemző az 16126-16136-16360 GVS1 motívum, amely Északkelet-Európa egyes populációiban is megtalálható.

|
Annino falu, 1917, dédnagymamám.
A teljes genomelemzés kimutatta, hogy az orosz és a mansi mtDNS-e különálló D5a3a klaszterbe egyesül, a koreaiak mtDNS-ét pedig külön ág képviseli. A teljes D5a3 haplocsoport evolúciós kora hozzávetőlegesen 20 ezer év (20560 ± 5935), míg a D5a3a mtDNS vonalak divergenciája megközelítőleg 5 ezer évnek (5140 ± 1150) felel meg. A D5 kifejezetten kelet-ázsiai csoport.
Szibériában a D4-es változatok abszolút túlsúlyban vannak. A D5 Japánban, Koreában és Dél-Kínában a legtöbb és legváltozatosabb. A szibériai népek közül a D5 sokféleségét és egyedi, tisztán etnikai változatainak jelenlétét a keleti mongol nyelvű csoportok, köztük a mongolizált evenkok körében figyelték meg. A D5a3 archaikus változata Koreában található, pontosabb elemzés szerint a D5a3a kora akár 3000 év is lehet, de a szülő D5a3 nagyon ősi, valószínűleg a mezolitikum.
Cserepovets, 1940
A rendelkezésre álló adatok alapján logikusnak tűnik a D5a3 eredete valahol a Távol-Keleten (Mongólia és Korea között), illetve Dél-Szibérián keresztül történő nyugati vándorlása. Valószínűleg úgy háromezer éve kerültek Európába közvetlen női felmenőim, akik Finnországban, Koreában, a helyi finnugor népeknél: számiknál, karjaliknál és vepszeneknél vetettek gyökeret. A Krivichivel keveredve ezek a haplocsoportok Vologda és a Novgorod régió modern lakosaihoz szálltak át.